

 下载:
下载:

-
肝内胆管癌(intrahepatic cholangiocarcinoma,ICC)是源于肝内胆管上皮细胞的高致死性恶性肿瘤[1],属于第二大类常见的原发性肝恶性肿瘤[2],约占所有肝恶性肿瘤的5%~20%[3]。ICC的发病率在全球范围内上升[4],中位生存期仅为11~13个月,5年生存率不足11%[5]。ICC手术治疗疗效不佳,仅20%~30%的患者有手术指征[6],术后5年生存率为20%~35%[7]。吉西他滨(gemcitabine)和顺铂(cisplatin)双药联合的一线化疗方案常伴随显著的不良反应[8]。虽然靶向药物如受体酪氨酸激酶抑制剂、异柠檬酸脱氢酶和成纤维细胞生长因子受体抑制剂有效但在数月内易出现耐药[9],提高ICC生存率并不明显[8]。因此,探索新型抗ICC药物、克服现有治疗方案的局限性并改善患者预后,已成为当前研究的迫切任务。
近年来,患者来源类器官(patient-derived organoid,PDO)技术为肿瘤精准医疗以及天然产物筛选提供了新工具[10]。相较于传统细胞系,类器官是由多种细胞类型组成的三维结构,能够模拟肿瘤在体内的组织结构,在保留患者个体特性方面具有显著性优势[11],已成为连接细胞实验与动物实验、基础研究与临床转化不可或缺的桥梁。研究表明,PDO模型可有效预测患者对药物的临床响应,为肿瘤个性化治疗提供依据,目前已经被广泛应用于肝癌、乳腺癌等多种实体瘤的研究[12]。运用PDO技术可完成活性化合物的药效评价,并且其测试结果具有更高的临床相关性,可加速抗ICC药物研发进程。
海洋微生物处在低温、低营养的极端环境中[13],其沉默基因会被激活,并通过代谢适应机制合成结构独特、活性显著的次级代谢产物,具有显著的抗菌、抗肿瘤、抗病毒活性[14]。从海洋放线菌Salinispora tropica发酵物中分离得到的沙利诺斯孢酰胺A(marizomib)已完成Ⅲ期临床试验,用于治疗胶质母细胞瘤[15],表明海洋微生物次级代谢产物在抗肿瘤药物研发中具有一定的潜力。本研究利用ICC的PDO模型对课题组自主采集与分离的极地海绵共附生链霉菌Streptomyces diastatius Y1来源的次级代谢产物2-庚基-3-羟基-4(1H)-喹诺酮[2-heptyl-3-hydroxy-4(1H)-quinolone,PQS]进行抗ICC活性评价,并用RBE细胞初步研究其作用机制,以期为研发抗ICC新药提供参考。
-
ICC组织样本取自2025年10月至2025年12月于上海交通大学医学院附属仁济医院(我院)接受手术的6例患者。纳入标准:①年龄≥18岁;②所有病例均经病理科确诊;③术前未进行任何治疗。排除标准:①合并其他恶性肿瘤或者已经发生肝转移患者;②经过多次系统治疗患者,记录患者的人口统计学特征、临床特点。本研究的实验方案经上海交通大学医学院附属仁济医院伦理委员会的审查并获得批准(批准号KY2025-158-B),所有样本的病理分析均在我院病理科进行。
-
人肝胆管癌RBE细胞由我院胆胰外科刘颖斌教授提供。化合物PQS由本课题组在极地放线菌链霉菌S. diastaticus Y1中分离、提取和制备,存放于我院药学部。CCK-8试剂盒(C6050,苏州新赛美生物科技有限公司);胎牛血清(10099141C)、DMEM培养基(C11995500CP)(Gibco公司);Annexin V-FITC/PI细胞凋亡检测试剂盒(AD10,日本同仁公司);肿瘤类器官基础培养基(B2131152)、组织保存液(K601005)、肿瘤组织消化液(K601003)、红细胞裂解液(E238010)、类器官冻存液(E238023)、类器官回收防粘连润洗液(E238002)、类器官传代消化液(E238001)、ICC完全培养基(K2104-LB)、细胞活性ATP检测试剂盒(E238003)(苏州bioGenous公司);基质胶(356234,美国Corning公司);顺铂(232120,Merck公司);S-vision免疫组化试剂盒含免疫组化法一抗(CYP3a蛋白兔多克隆抗体,GB115301-100)、二抗(G1312-100T)(赛维尔生物科技有限公司);碘化丙啶/RNase染色缓冲液(550825,BD Pharmingen公司);核因子κB p65抗体(8242S)、核因子κB p50抗体(13586S)、细胞周期蛋白依赖性激酶2(cyclin-dependent kinase 2,CDK2)抗体(18048S)、CDK4抗体(12790S)、β-微管蛋白抗体(2146S)、HRP标记山羊抗兔IgG二抗(7074S)(CST公司)。
-
生物安全柜(HR30-IIA2)、二氧化碳培养箱(371)、低温高速离心机(Micro 21R)、流式细胞仪(Attune Nxt)、多功能酶标仪(VariousKan LUX)(美国Thermo Fisher Scientific公司);普通倒置显微镜(TS100,日本Nikon公司)。
-
来源于肝内胆管癌患者的肿瘤组织样本在无菌手术室取得,用无菌PBS清洗后保存于预冷组织保存液中运至实验室。用含有青霉素和链霉素的肿瘤基础培养基洗涤,并剔除周围脂肪、多余纤维组织和血。用手术剪将肿瘤组织剪碎成0.5~1.0 mm3的小块,浸没于肿瘤组织消化液,于37℃恒温摇床消化30 min。待组织能够通过1 ml移液头时,加入含10%胎牛血清的培养基终止消化。经70 μm滤网过滤获得单细胞悬液,细胞悬液以5×105/ml密度与基质胶以3∶7的体积比于冰上混匀;取50 μl 接种至 24孔培养板,37℃固化15 min,每孔加入500 μl ICC的完全培养基进行维持培养,每隔2 d在显微镜下进行观察。PDO模型构建成功的标志为形成具有明显腔隙的囊泡样结构且边界清晰。
-
待基质胶中大部分的PDO长至200 μm克隆大小时,即可进行传代。传代时需用类器官回收防黏连润洗液对移液头和离心管进行润洗,以防PDO附着在移液头和离心管上而造成类器官损失,用200 μl移液头刮下孔板里PDO-基质胶复合物,并且添加200 μl类器官基础培养液,用200 μl移液头吹打复合物使PDO与基质胶分离。分离好的PDO用类器官传代消化液消化至单个细胞,用水平离心机以500×g转速离心3 min后按相同密度和相同条件重新接种培养。显微镜下观察PDO组织形态和生长状态,1~2 d换液1次,每4~5 d传代1次。为了保存PDO,将PDO冻存。刮下PDO-基质胶复合物,吹打使PDO与基质胶分离,离心去上清后用1 ml类器官冻存液重悬PDO,配制成PDO悬液,置于液氮中长期储存。
-
将冻存的PDO置于37℃水浴2 min,用预热的肿瘤基础培养基清洗1次,在水平离心机上以300×g转速离心3 min弃上清液,加入60 μl基质胶并轻柔混匀,然后将悬液接种至24孔板中,37℃细胞孵箱培养25 min待基质胶凝固后;每孔添加500 μl肿瘤类器官培养基,置37℃细胞孵箱培养,显微镜下观察PDO的生长状态。
-
收集患者ICC组织,将ICC组织和PDO置于4%多聚甲醛中固定,随后融化琼脂糖、ICC组织和PDO包埋固定、脱水、融蜡、透明化处理、浸蜡、蜡块包埋。最后参照流程进行HE染色,染色后脱水、封片、晾干,于显微镜下观察包括核形态、腺体结构及细胞异型性等组织形态学特征并拍照。
-
免疫组化法检测患者ICC组织和PDO组织中CYP3a蛋白的表达,包括组织脱蜡水化、细胞通透与封闭,抗原修复、血清封闭、孵育一抗 CYP3a(稀释比例 1∶500)4℃反应过夜、HRP 标记二抗(稀释比例 1∶1 000)室温孵育及切片显色等步骤。最后经脱水透明后封片,于倒置显微镜下观察组织CYP3a蛋白的阳性表达定位、显色强度及分布范围并采集图像进行分析。CYP3a蛋白阳性表达以DAB显色呈棕黄色颗粒为准,根据显色强度和阳性细胞比例进行半定量检测蛋白表达水平。
-
取稳定传代的PDO模型,经消化制备成含有1×104/ml单细胞悬液。将单细胞悬液与基质胶按照3∶7体积比混匀制成20 μl单细胞与基质胶混合物,接种至24孔板中,于37℃细胞培养箱固化25 min后放入培养箱开始计时,于24 h后加入PQS至培养基(PQS终浓度为0、2.5、5、10 μmol/L),另加入顺铂作为阳性对照药(10 μmol/L),并且对在培养箱培养24 h后的PDO形态进行拍照作为PQS作用0 h的对照。PQS持续作用96 h,每隔24 h进行拍照记录,观察细胞是否萎缩死亡,并进行PDO计数评价PQS对PDO生长的影响。
再取稳定传代的PDO模型,经消化制备成含有1×103/ml单细胞悬液,将单细胞悬液与基质胶按照3∶7体积比混匀制成10 μl单细胞与基质胶混合物,接种至96孔板中,于37℃细胞培养箱固化25 min后培养24 h;放入培养箱开始计时,于24 h后加入PQS至培养基(PQS终浓度为0、1.25、2.5、5、10、20 μmol/L),另加入顺铂作为阳性对照药(10 μmol/L)。药物持续作用96 h后采用ATP法检测细胞活力,通过拟合浓度-响应曲线评估PQS对PDO生长的抑制活性。
-
人肝内胆管癌细胞系RBE细胞用1640+10%FBS+1%青霉素链霉素的培养基于37℃、5% CO2培养箱内细胞贴壁生长,确认无支原体污染,复苏后传代不超过30次。
-
取对数生长的RBE细胞,显微镜下观察细胞形态是否良好。将细胞进行消化、终止、离心、重悬制成1×104/ml的细胞悬液,接种于在96孔板中,接种体积为100 μl。24 h贴壁后每孔加入10 μl不同浓度的PQS溶液,其PQS终浓度分别为20、10、5、2.5、1.25、0.65、0.325 μmol/L,同时设10 μmol/L顺铂阳性药物组,每个浓度设复孔3个,细胞对照组(含细胞但不含药物)、空白对照组(仅有培养基无细胞)。将培养板放入37℃、5% CO2培养箱内培养48 h。每孔加入10 μl的CCK-8试剂,避光孵育30 min,在酶标仪450 nm波长进行检测。检测过程中使细胞对照组吸光度值在0.8~1.2。细胞存活率(%)=(A给药组−A空白对照组)/(A细胞对照组−A空白对照组)×100%),用Graphpad模拟非线性曲线回归进行曲线拟合,确定后续实验最佳给药剂量。
-
取对数生长的RBE细胞,将RBE细胞消化离心成细胞悬液,按200/孔接种于12孔板中,将贴壁后细胞分为细胞对照组以及细胞给药组,在细胞对照组中加入DMSO、在细胞给药组中加入PQS至培养基(PQS终浓度为1、2、4 μmol/L),持续培养10~14 d,期间每2 d换药1次。待克隆肉眼可见后,用4%多聚甲醛固定15 min,0.1%结晶紫染色10 min,PBS清洗后晾干拍照。以≥50个细胞为克隆的标准计数,使用ImageJ软件计数克隆数并进行统计分析。
-
取对数生长的RBE细胞,将RBE细胞消化离心成细胞悬液,按1×105/每孔细胞接种于6孔板中,将贴壁后细胞分为细胞对照组以及细胞给药组,在细胞对照组中加入DMSO、在细胞给药组中加入PQS至培养基(PQS终浓度为1、2、4 μmol/L)处理48 h,收集贴壁细胞,用1 ml PBS进行重悬细胞,后将细胞逐滴加入装有3 ml的无水乙醇5 ml离心管中,在−20℃冰箱固定12 h。固定好的细胞用水平离心机以
1500 r/min离心5 min;弃上清,将细胞用PBS重悬洗涤后用水平离心机以1000 r/min离心3 min;弃上清液,加入400 µl碘化丙啶/RNA酶染色缓冲液重悬细胞,在37℃培养箱中避光孵育20 min。孵育结束后以1000 r/min离心3 min,弃染色缓冲液,于细胞沉淀中加入PBS离心洗涤,重悬细胞,用流式细胞术检测细胞周期。 -
取对数生长的RBE细胞,将RBE细胞消化、离心,制备成细胞悬液,按1×105 /每孔细胞接种于6孔板中,将贴壁后细胞分为细胞对照组以及细胞给药组,在细胞对照组中加入DMSO、在细胞给药组中加入PQS至培养基(PQS终浓度1、2、4 μmol/L)处理48 h,收集贴壁及漂浮细胞,按照Annexin V-FITC/PI试剂盒说明书染色。37℃烘箱孵育抗体,结束后洗涤2次,立即使用流式细胞仪检测凋亡率,用FlowJo软件分析数据。
-
取对数生长的RBE细胞,将RBE细胞消化、离心,制备成细胞悬液,按1×105/每孔细胞接种于6孔板中,将贴壁后细胞分为细胞对照组以及细胞给药组,在细胞对照组中加入DMSO、在细胞给药组中加入PQS至培养基(PQS终浓度为1、2、4 μmol/L)处理48 h,收集贴壁及漂浮细胞,提细胞总蛋白,采用BCA蛋白检测试剂盒检测蛋白浓度。取12 µg蛋白样品经十二烷基硫酸钠-聚丙烯酰胺电泳分离后,冰浴湿转1 h至PVDF膜上;用5%脱脂牛奶封闭40 min后,分别加入核因子κB p65抗体、核因子κB p50抗体、CDK2抗体、CDK4抗体、β-微管蛋白抗体孵育过夜;然后用TBST缓冲溶液清洗3次,每次10 min;室温孵育山羊抗兔IgG二抗HRP标记40 min后,用TBST缓冲溶液清洗3次,每次10 min。用ECL化学发光液反应,并于G-BOX成像分析系统获得蛋白免疫印记图。采用ImageJ软件对图像进行分析,以β-微管蛋白为内参,以目标蛋白与内参蛋白条带积分吸光度比值表示目标蛋白的表达水平。
-
采用GraphPad Prism 7.0软件对数据进行统计分析,用平均值±标准方差($\bar x $±s)表示,两组间均数采用t检验比较,多组间均数采用One-Way ANOVA检验。
-
从6例ICC组织样本中,成功构建了2例PDO模型,如表1所示。所有组织运输时间均小于60 min,起始的活细胞数≥1×106。
表 1 所构建患者来源类器官(PDO)的患者临床信息
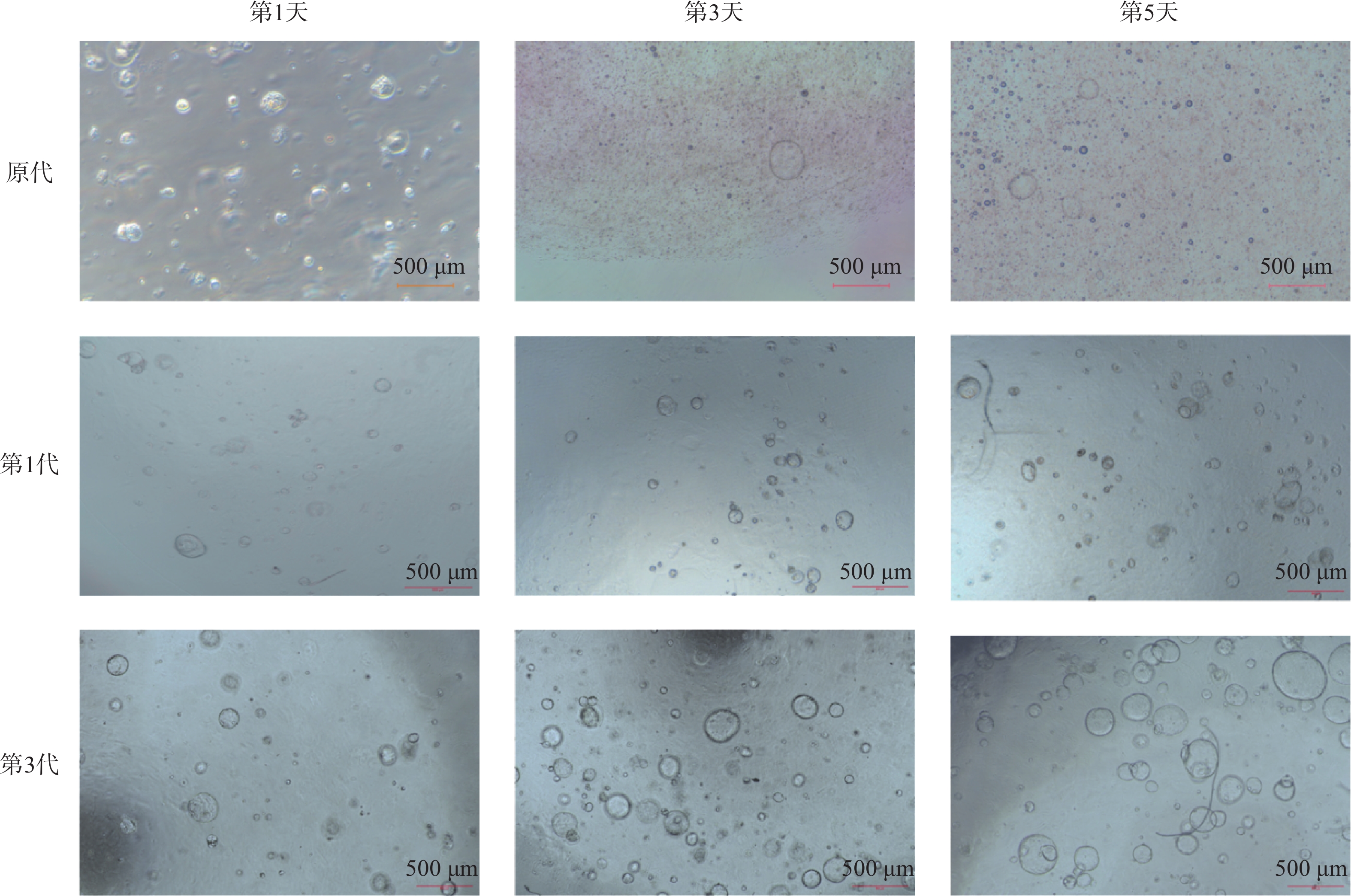
PDO编号 性别 年龄/岁 肿瘤类型 pT pN pM 分期 PDO#1 男 48 ICC T1 N0 M0 Ⅰ PDO#2 女 74 ICC T2 N1 M0 ⅢB ICC:肝内胆管癌;pT:病理T分期;pN:病理N分期;pM:病理M分期;T1:肿瘤直径≤2cm,无血管侵犯;T2:肿瘤直径>2cm,或伴有血管侵犯;N0:无区域淋巴结转移;N1:有区域淋巴结转移;M0:无远处转移;Ⅰ期 / ⅢB期根据TNM分期系统诊断。 2名患者的PDO呈现的形态大致相同。原代构建第1天铺板时细胞数多,其中含有肝纤维细胞、ICC细胞,肝细胞癌细胞。但是第3天以及第5天只有具有干性的ICC细胞能够在基质胶以及在ICC培养基条件下进行增殖,其余细胞则不能够在此条件下进行生长。PDO呈现典型三维囊泡样球形,直径50 μm,连续传代2代后形态保持稳定,背景干净,数量明显增多,直径可达100 ~ 300 μm,如图1所示。此时的PDO能够稳定传代,传代次数可达20代次。前几代PDO活力强可将其冻存。
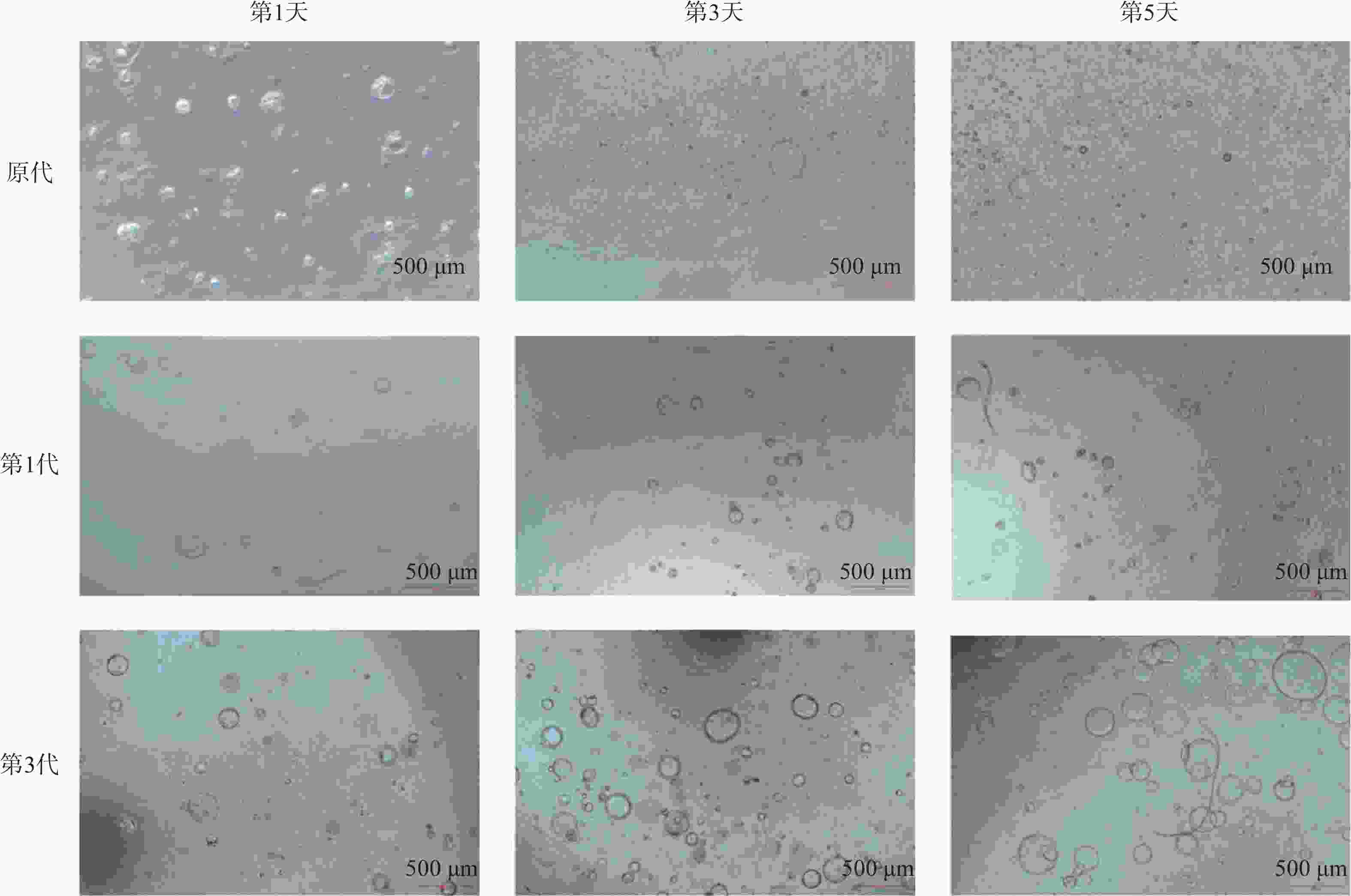
图 1 PDO#1培养体系的动态形态学观察(比例尺=500 µm)
-
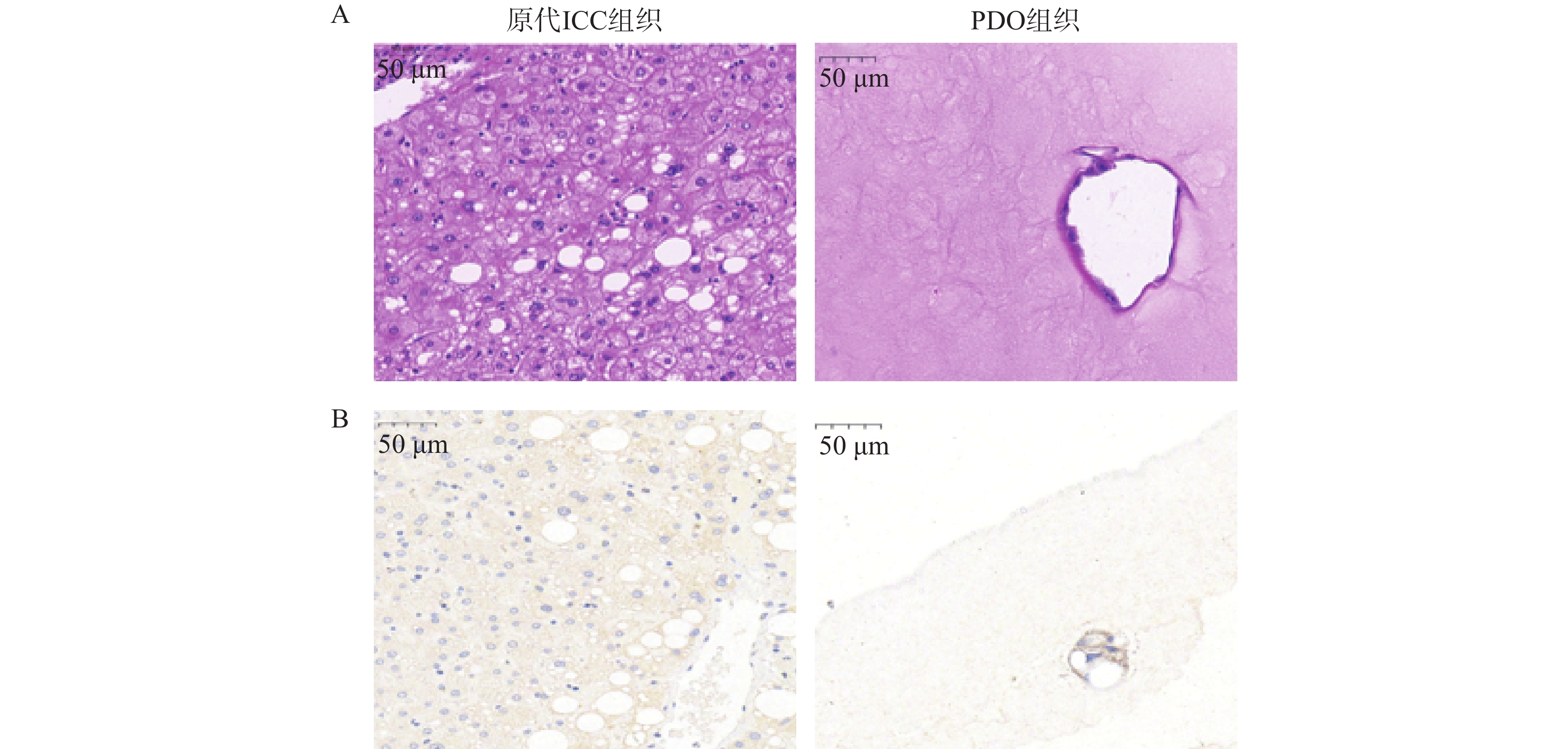
PDO构建成功的标志主要可以从2个层面进行评价。形态学层面在明场具有空心的结构,HE染色显示的组织形态与原始肿瘤组织高度一致。肿瘤标志物层面,通过免疫组化确认其表达胆管上皮谱系的特异性蛋白[16]。本研究选择了形态学层面以及肿瘤标志物层面进行鉴定。如图2A的HE染色结果显示,原代ICC组织中具有空腔结构,PDO组织形态表现为单层不规则腺腔,核大深染、核仁明显,腔缘可见微绒毛及乳头状内折,细胞异型性显著,与原发灶形态学特征基本一致。图2B所示,通过免疫组化检测ICC标志物CYP3a表达,发现PDO组织与原代ICC组织均表达CYP3a蛋白,并且与原代ICC组织形态保持高度相似性。
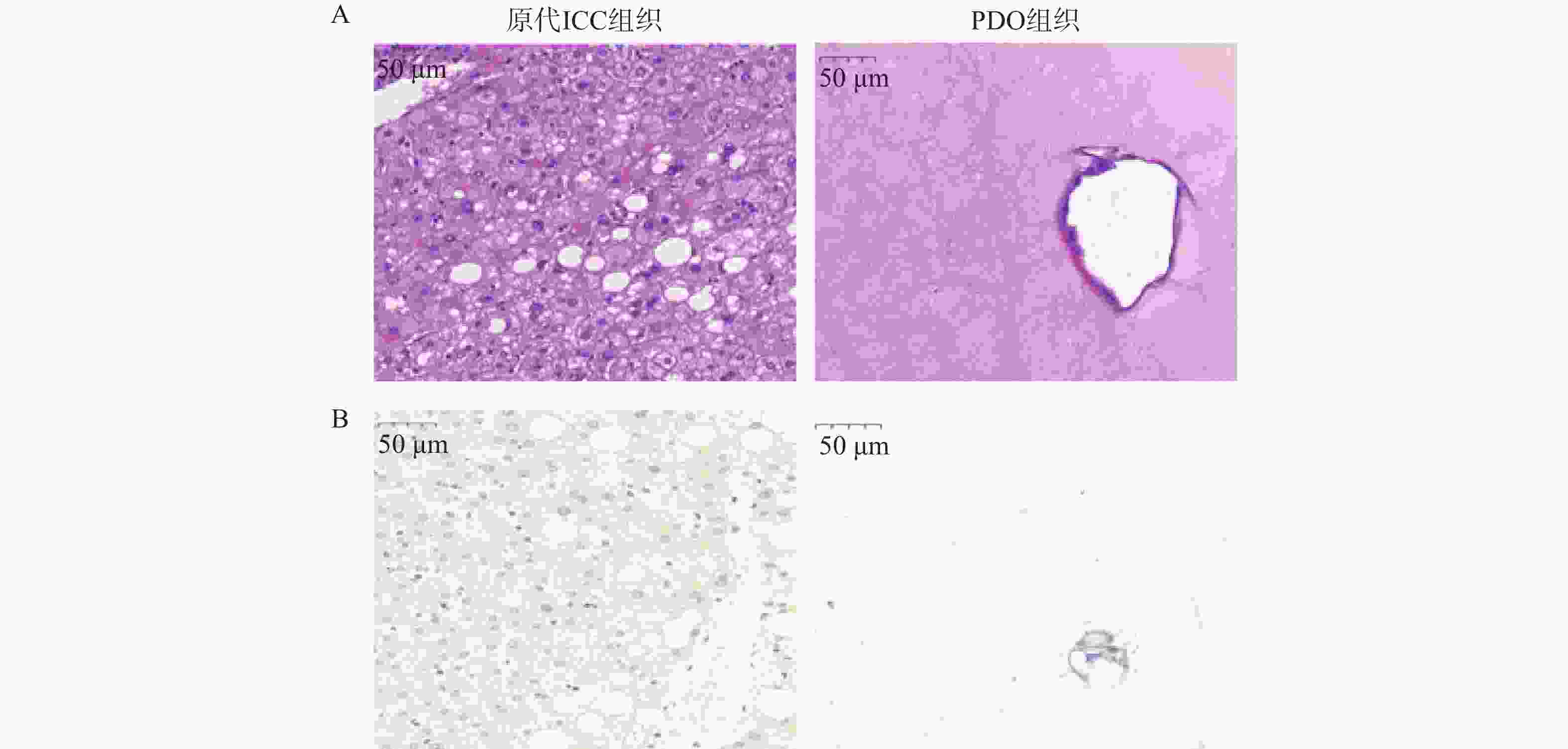
图 2 PDO组织与原代ICC组织形态表征(比例尺=50 µm)
-
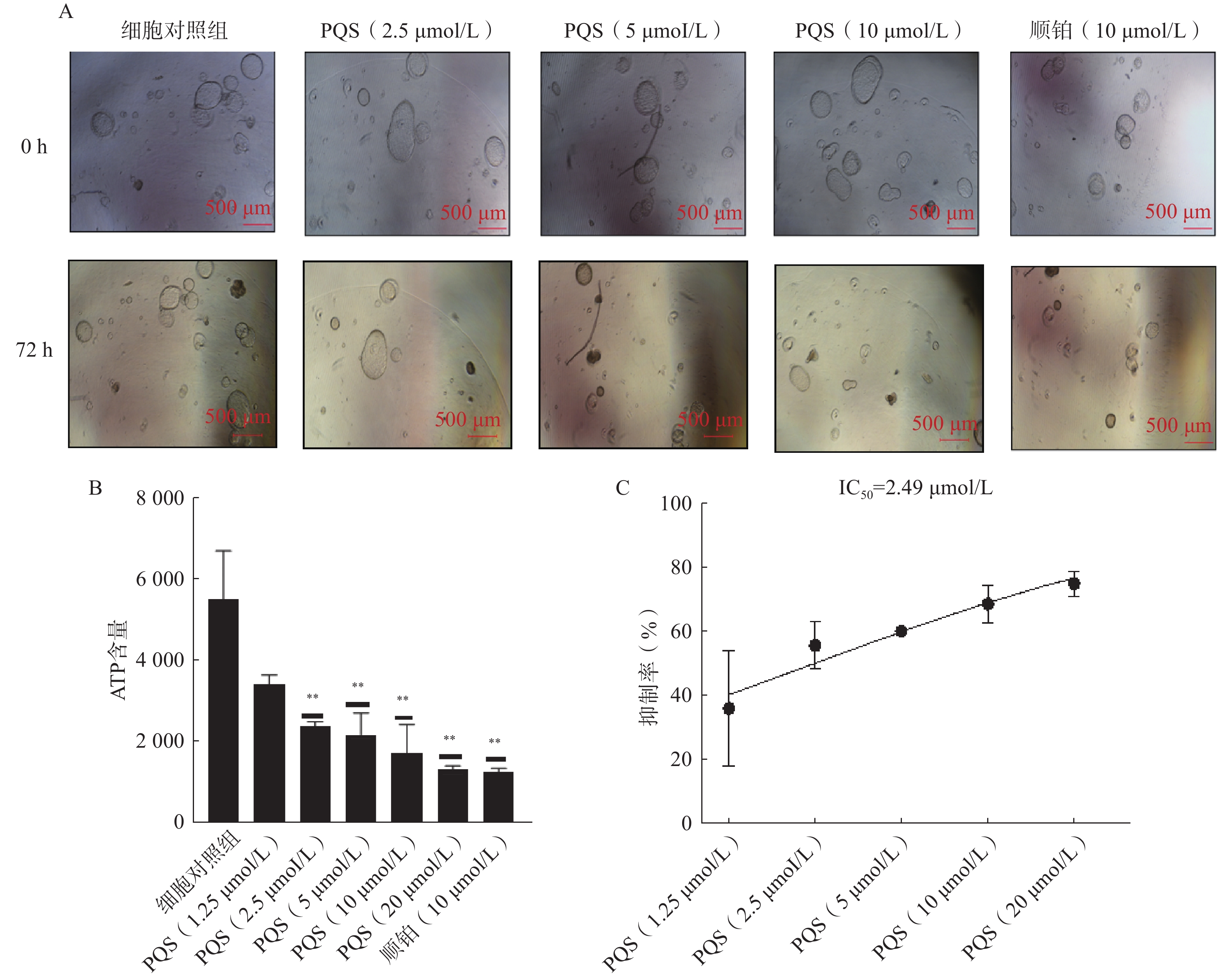
在显微镜下观察同一个视野下铺板贴壁后细胞对照组及不同浓度(2.5、5、10 μmol/L)PQS处理0和72 h后PDO形态变化。结果如图3A所示,细胞对照组PDO能够正常生长,PDO数量增多;顺铂组PDO数量减少,部分PDO萎缩死亡;PQS在2.5、5、10 μmol/L下可浓度依赖地减少PDO数量。
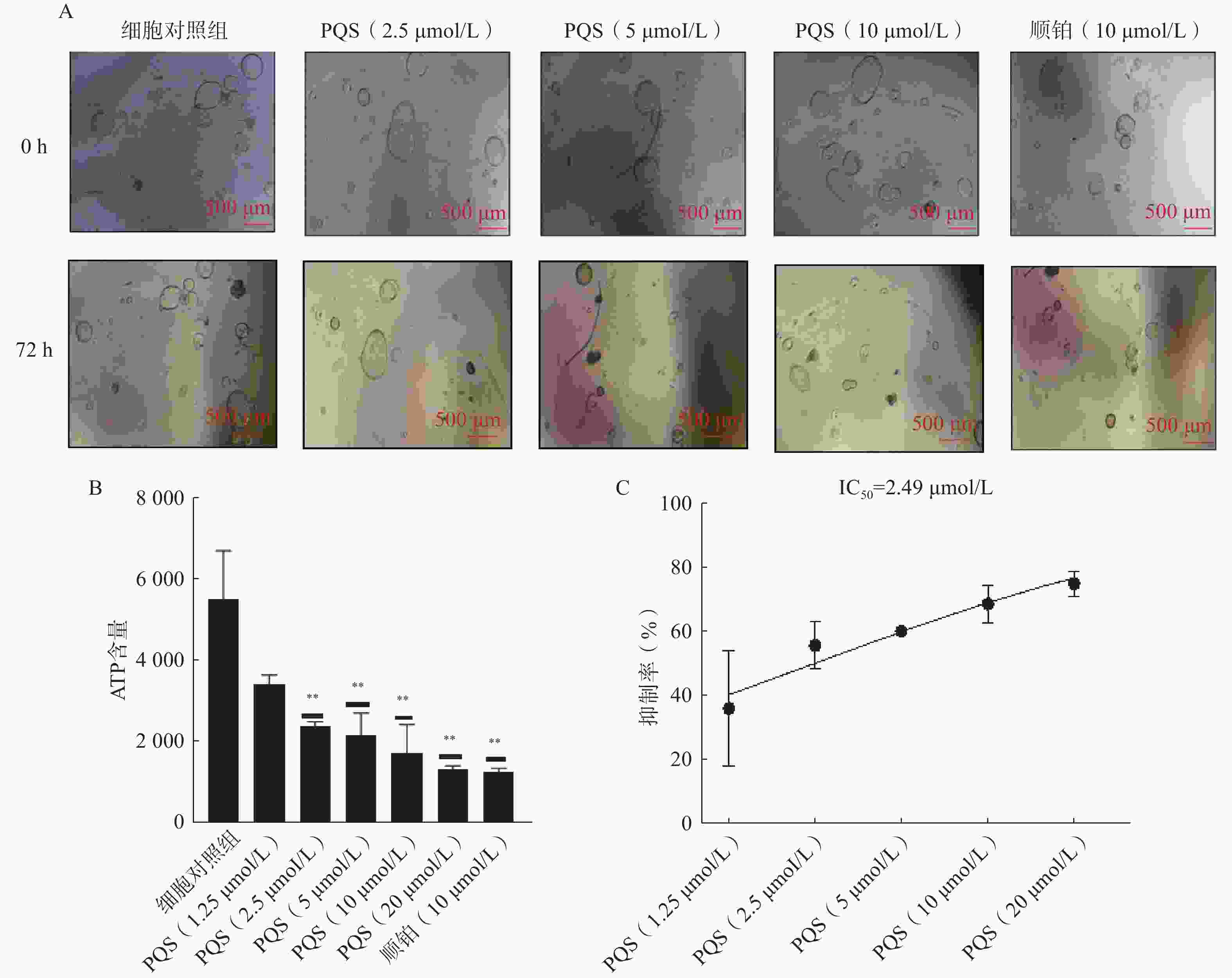
图 3 PQS对PDO生长的抑制活性(比例尺=500 µm,$ \bar{x} $±s,n=3)
通过ATP法分析PQS抑制PDO生长的浓度反应, 结果如图3B所示,PQS对PDO生长的半数抑制率(IC50)为2.49 µmol/L。图3C所示,顺铂及不同浓度(2.5、5、10 μmol/L)PQS处理能够具有浓度依赖性地减少PDO的ATP含量。以上结果表明,PQS具有抗ICC活性。
-
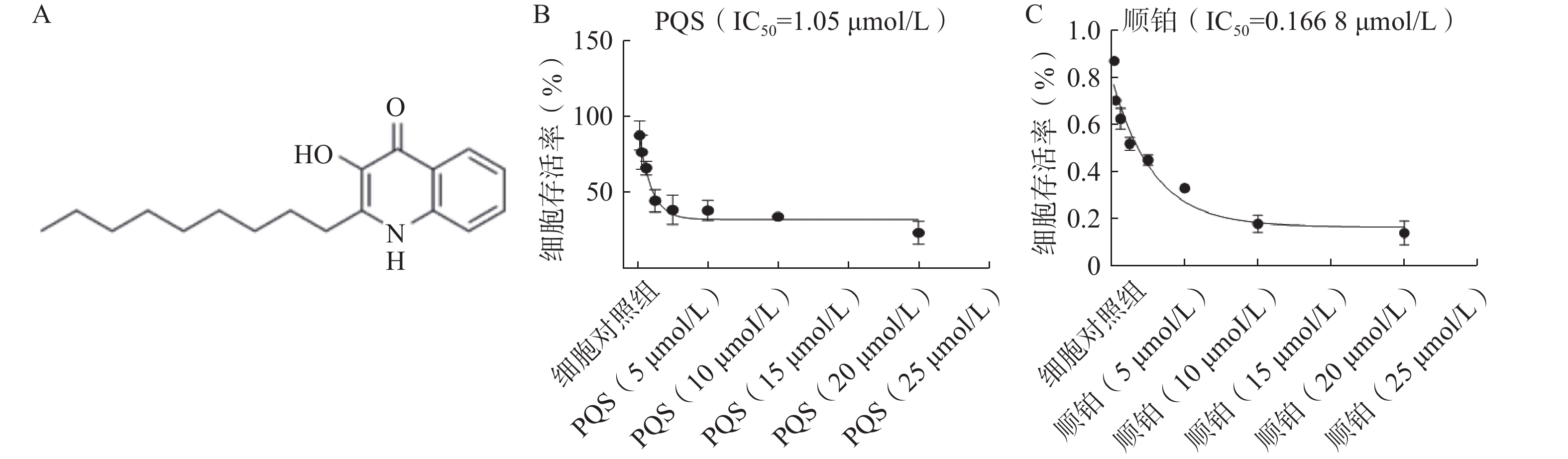
PQS的化学结构如图4A所示。如图4B所示,PQS(20、10、5 µmol/L)处理48 h可浓度依赖地降低RBE细胞的存活率,其IC50值为1.05 µmol/L。如图4C所示,顺铂处理48 h细胞存活率同样随着顺铂浓度增加而降低。参考以上结果,确定1、2、4 µmol/L作为PQS的给药浓度,并选择处理48 h进行后续实验。
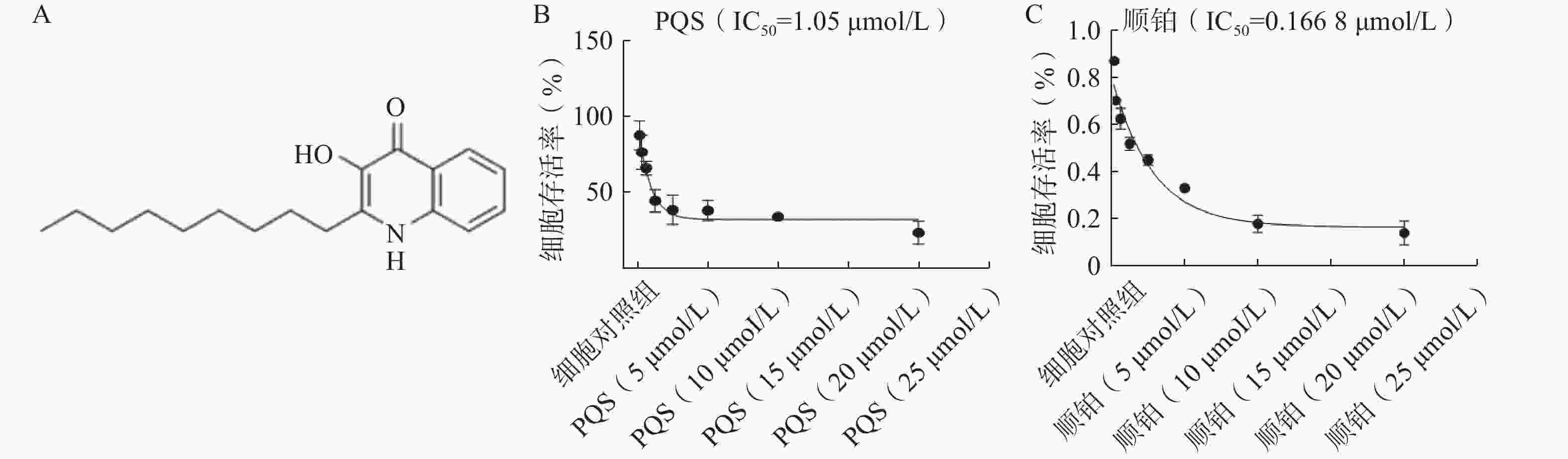
图 4 PQS结构及其对RBE细胞存活率的影响($\bar x $±s, n=3)
-
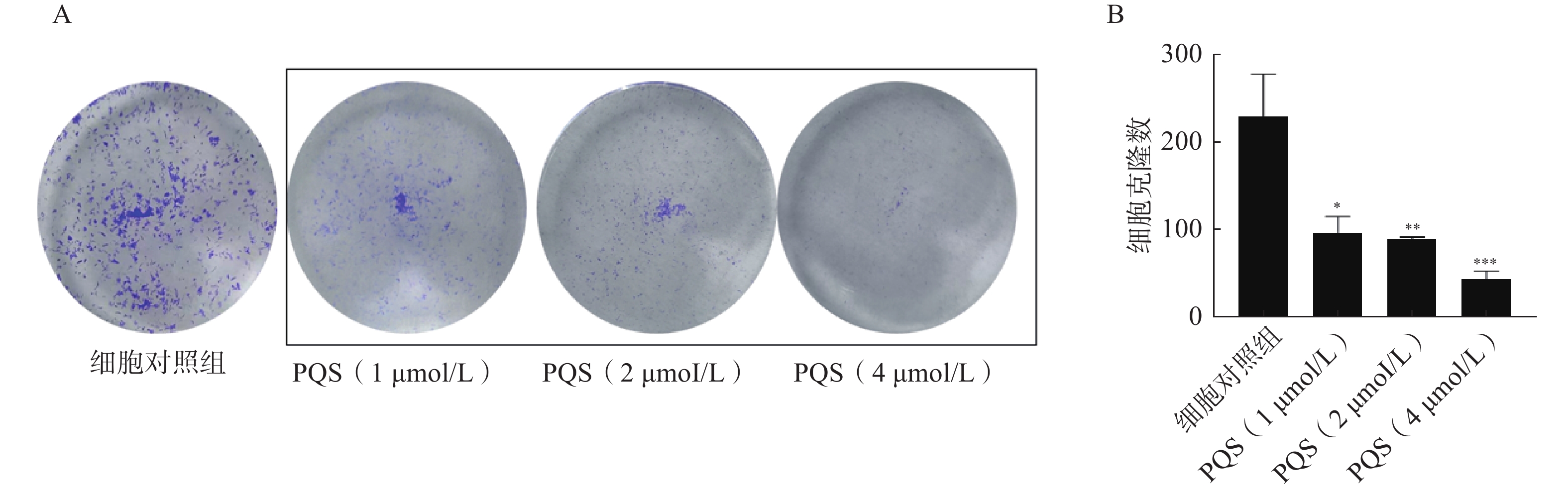
如图5所示,与细胞对照组比较,PQS(1、2、4 µmol/L)给药组细胞克隆形成数均显著降低(P<0.001),且呈浓度相关性,表明PQS能够抑制RBE细胞的集落形成能力,即抑制RBE细胞增殖。

图 5 PQS对RBE细胞集落形成的影响($\bar x $±s, n=3)
-
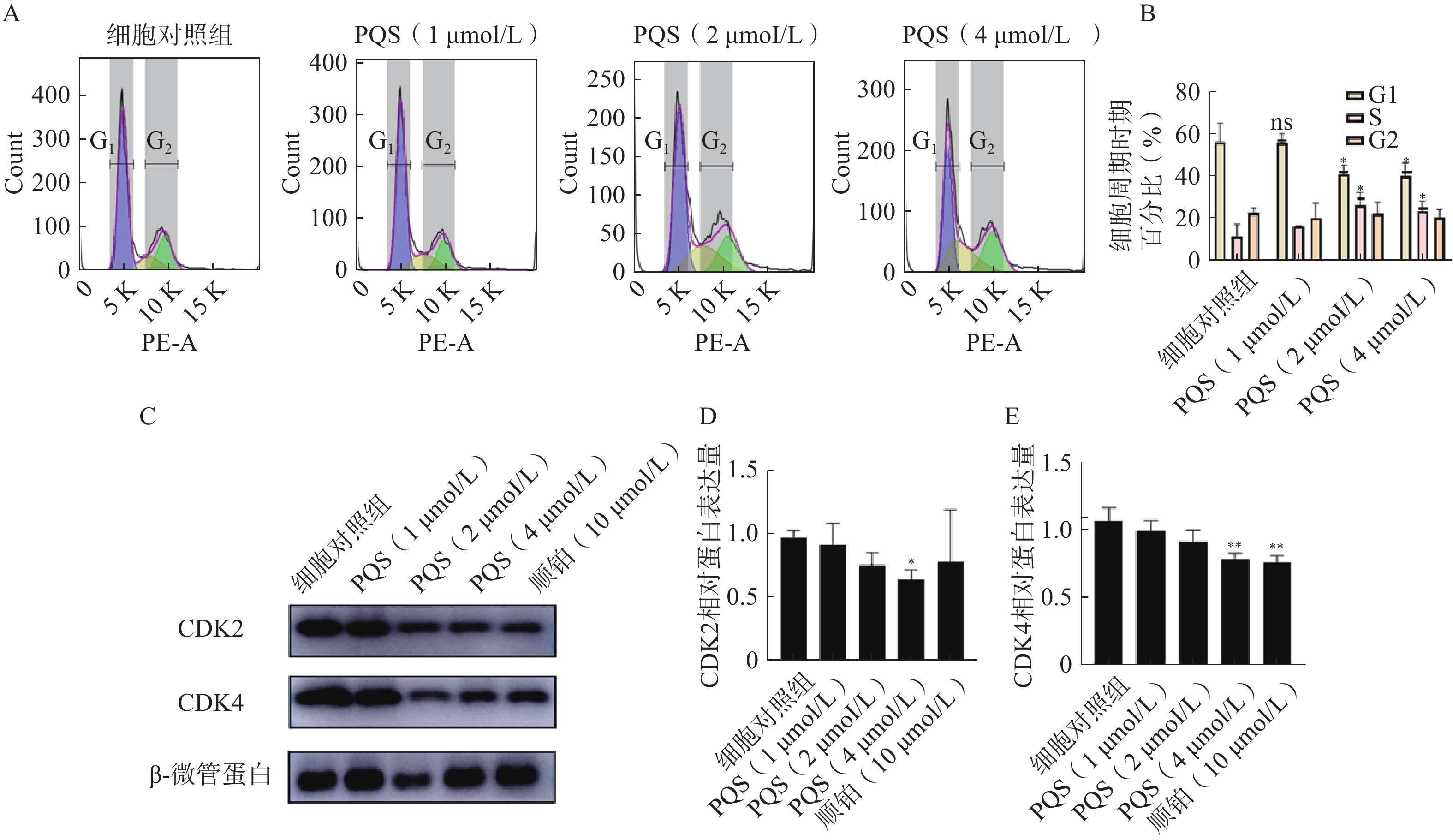
如图6A所示,PQS处理48 h后,随着PQS浓度的增加,处于G1期的RBE细胞百分比较对细胞照组显著减少,S期细胞百分比显著增加(P<0.05),表明PQS可将RBE细胞周期阻滞于S期。随后检测S期的特异性周期蛋白CDK2、CDK4的表达水平,结果如图6B所示,PQS组细胞中CDK2、CDK4的表达水平较细胞对照组均有所降低,且高浓度(4 µmol/L)更为显著(P<0.05,P<0.01)。
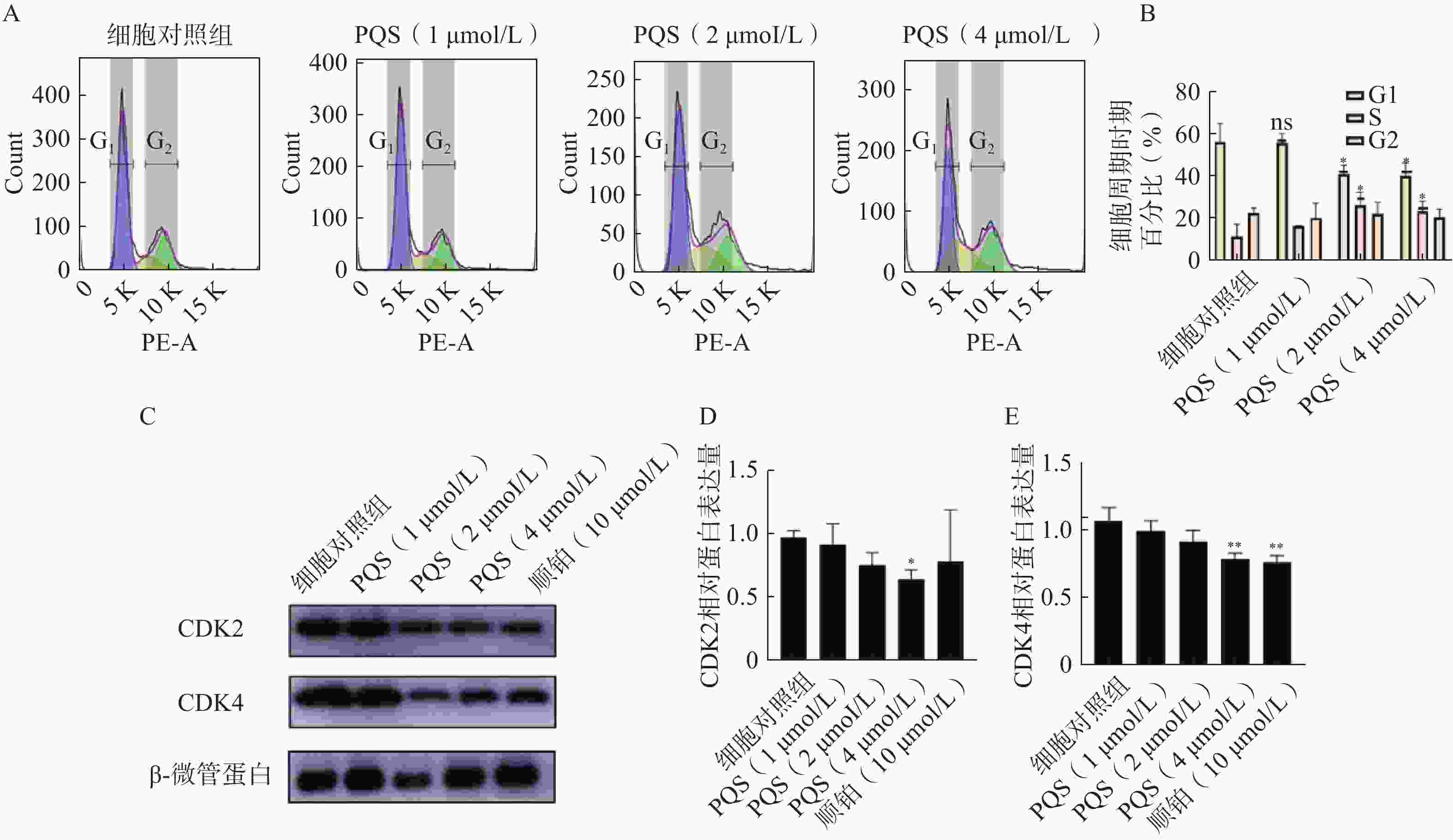
图 6 PQS对RBE细胞周期和细胞周期蛋白表达的影响($\bar x $±s, n=3)
-
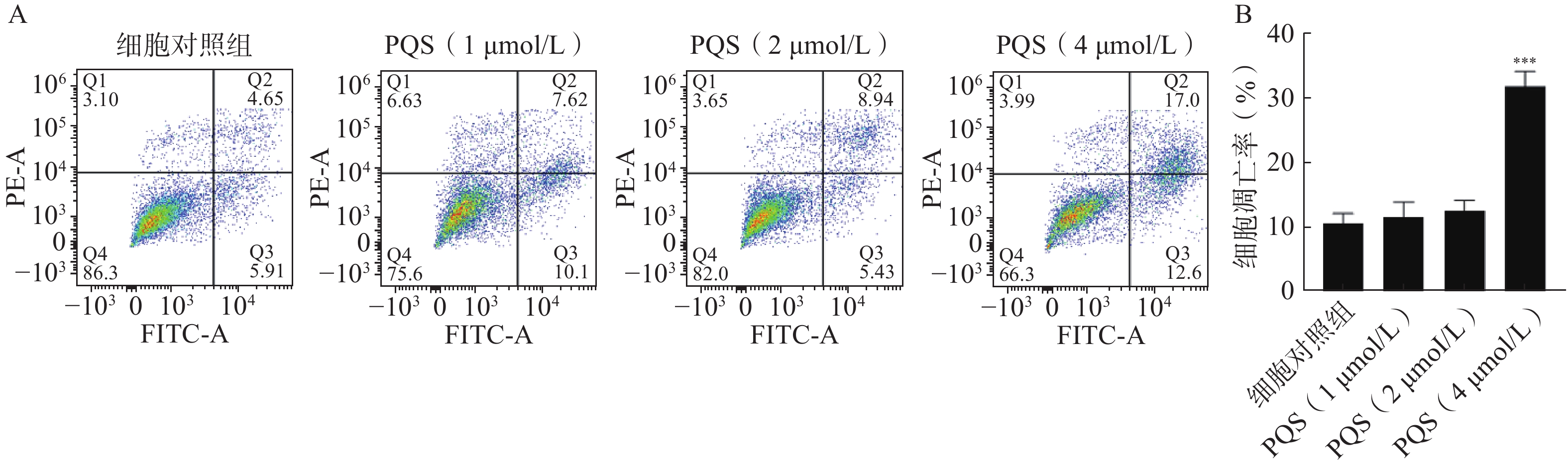
如图7所示,流式细胞术分析表明,PQS处理细胞48 h后与细胞对照组相比,PQS显著提高RBE细胞凋亡率(P<0.01),说明其具有诱导RBE细胞凋亡的作用。
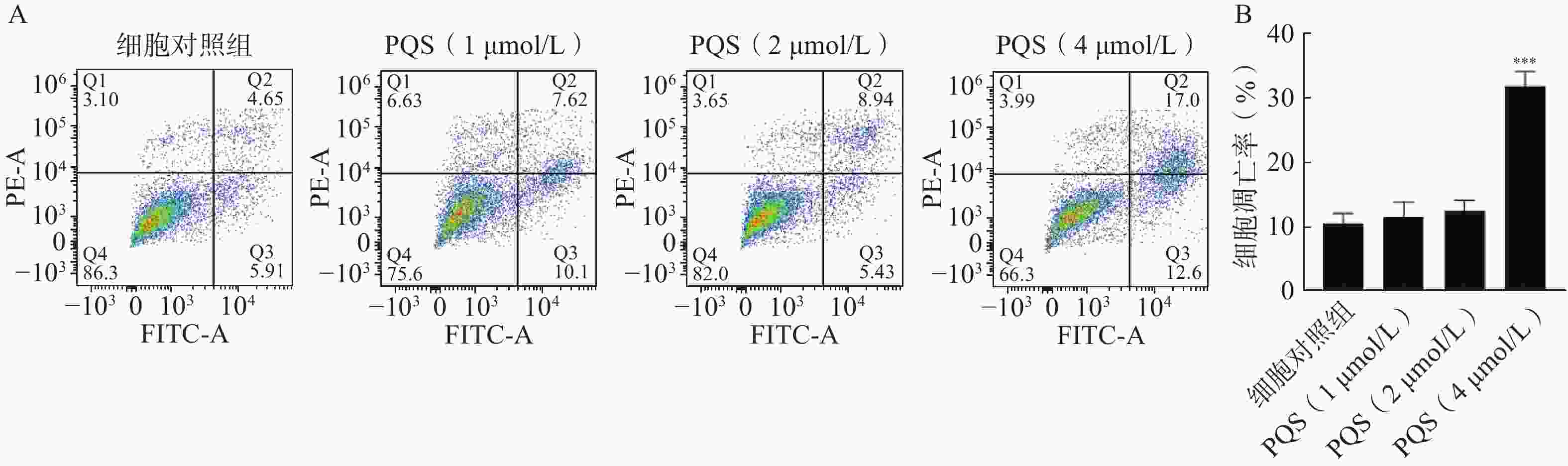
图 7 PQS对RBE细胞凋亡的影响($\bar x $±s, n=3)
-
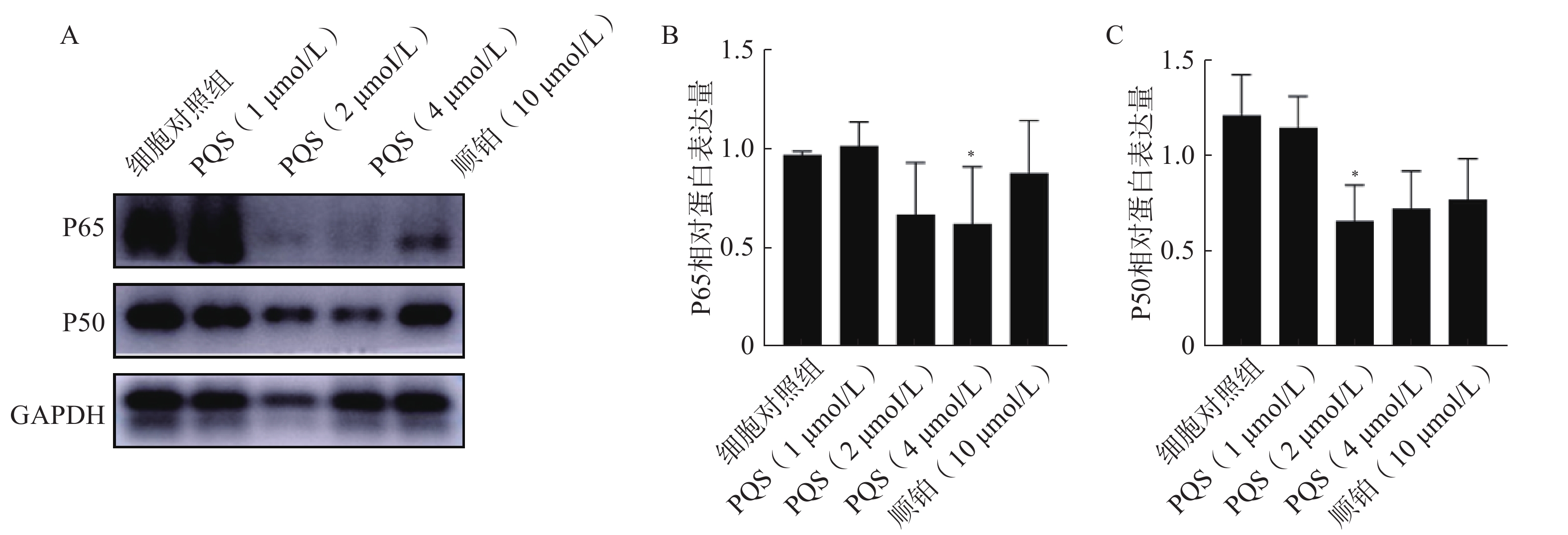
如图8所示,PQS处理细胞48 h后,与细胞对照组相比,PQS处理组P65、P50蛋白表达水平均显著降低(P<0.05),表明PQS抑制RBE细胞中NF-κB信号通路的活化。
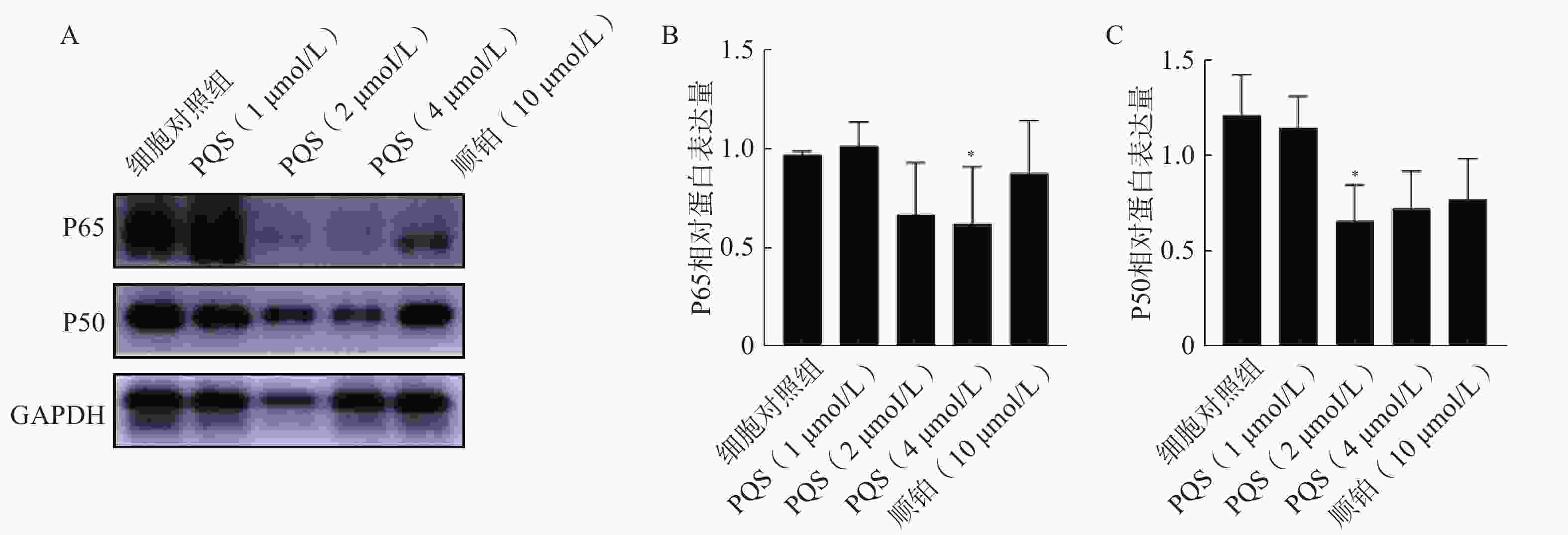
图 8 PQS对RBE细胞中P65、P50蛋白表达的影响($\bar x $±s, n=3)
-
PQS是一种结构独特且具有多方面药理活性的极地海绵放线菌次级代谢产物,包括抗肿瘤活性以及神经保护作用[17]。在抗肿瘤活性方面,有研究报道PQS对霍奇金淋巴瘤细胞具有抑制作用[18],但尚无研究探讨该化合物对ICC的潜在作用。此外,PQS已被证实可通过铁死亡途径诱导巨噬细胞死亡[19],且大量文献报道其与先天免疫调控存在关联[20]。
本研究成功建立了ICC患者来源的肿瘤类器官,通过组织形态学与组织病理学鉴定表明,其三维结构与病理标志物与原代ICC一致。通过构建的PDO模型,发现PQS能够有效抑制PDO增殖,具有显著的抗ICC活性。随后在RBE细胞模型上初步探讨了其抗ICC机制,发现PQS可浓度依赖性抑制RBE细胞克隆形成,将RBE细胞阻滞在S期,而且可诱导RBE细胞凋亡,该作用与NF-κB通路相关。
综上所述,本研究成功构建了ICC PDO模型,保留了原代ICC的组织学特征,而且该技术所需样本量较少,恰好与海洋天然产物的稀缺性形成互补,为海洋来源抗肿瘤活性成分的精准高效验证提供了技术可行性。但PDO模型也具有一定的局限性,其缺乏体内微环境,且样本来源较为单一,未能充分涵盖ICC的分子亚型异质性。
本研究在类器官和细胞水平发现,PQS具有抗ICC的活性,提示PQS具有作为潜在抗ICC先导分子应用潜力。但本研究尚未明确PQS的直接作用靶标,对其分子机制尚待深入探讨;而且尚未进行体内动物实验验证,其体内药效、药代动力学特征及安全性评价仍有待进一步考察。
Evaluation of anticancer activity of marine microbial secondary metabolites based on intrahepatic cholangiocarcinoma organoid models and study on its induction of cancer cell apoptosis
-
摘要:
目的 探讨极地海洋微生物次级代谢产物2-庚基-3-羟基-4(1H)-喹诺酮(PQS)的抗肝内胆管癌(ICC)活性和机制。 方法 选用来自上海交通大学医学院附属仁济医院的ICC肿瘤组织构建患者来源类器官(PDO)模型;用HE染色检测患者ICC组织和PDO组织的形态;用免疫组化法检测CYP3a在患者ICC组织和PDO组织的表达。用三磷酸腺苷(ATP)法检测PQS对稳定传代PDO生长的抑制活性,拟合剂量-反应曲线计算IC50,评价PQS的抗ICC活性。在人肝胆管癌RBE细胞模型中,用CCK-8法评价PQS对RBE细胞增殖活性的影响,平板克隆法检测对RBE细胞集落形成能力的影响,流式细胞术检测对RBE细胞周期与凋亡的影响,Western蛋白印迹法检测对RBE细胞周期蛋白依赖激酶2(CDK2)、CDK4、RelA蛋白(P65)、核因子κB1(P50)蛋白表达的影响。 结果 成功建立2例ICC PDO,组织形态学观察发现,连续传代后的PDO组织与患者ICC组织形态基本一致,均为囊泡样结构。免疫组化法检测结果表明,PDO组织与患者ICC组织均表达CYP3a; ATP法测定结果表明,PQS能够有效抑制PDO组织中ATP含量,拟合曲线得到IC50为2.49 μmol/L。在RBE细胞模型中,PQS可浓度依赖性抑制RBE细胞活力,拟合曲线得到IC50为1.05 μmol/L;同时PQS 1、2、4 μmol/L可抑制RBE细胞集落形成,将RBE细胞周期阻滞在S期,并诱导RBE细胞发生凋亡,下调NF-κB信号通路相关蛋白的表达。 结论 成功构建ICC PDO,证实PQS具有抗ICC活性。PQS可抑制RBE细胞增殖,可能通过NF-κB信号通路将细胞周期阻滞在S期,诱导RBE细胞发生凋亡。 Abstract:Objective To investigate the anti-intrahepatic cholangiocarcinoma (ICC) activity and mechanism of 2-heptyl-3-hydroxy-4(1H)-quinolone (PQS), a secondary metabolite of polar marine microorganisms. Methods Patient-derived organoid (PDO) models were established using intrahepatic cholangiocarcinoma (ICC) tumor tissues obtained from Renji Hospital, Shanghai Jiao Tong University School of Medicine. Hematoxylin and eosin (HE) staining was performed to assess the histomorphological characteristics of both patient ICC tissues and corresponding PDOs. Immunohistochemistry (IHC) was employed to evaluate CYP3a expression in patient ICC tissues and PDOs. The antiproliferative activity of PQS against stably passaged PDOs was determined using an adenosine triphosphate (ATP)-based bioluminescence assay, and dose-response curves were fitted to calculate the half-maximal inhibitory concentration (IC50) for assessing the anti-ICC efficacy of PQS. In the human intrahepatic cholangiocarcinoma RBE cell model, the effects of PQS on RBE cell proliferation were evaluated by the cell counting kit-8 (CCK-8) assay; colony formation capacity was assessed by the plate colony formation assay; cell cycle distribution and apoptosis were analyzed by flow cytometry; and the protein expression levels of cyclin-dependent kinase 2(CDK2), CDK4, RelA(p65), and nuclear factor-κB1(p50) were detected by Western blotting. Results Two ICC PDO models were successfully established. Histomorphological observation revealed that the PDO tissues after serial passaging exhibited morphological features essentially consistent with the corresponding patient ICC tissues, both presenting as cystic vesicle-like structures. Immunohistochemical analysis demonstrated that CYP3a was expressed in both PDO tissues and patient ICC tissues. ATP-based bioluminescence assay results indicated that PQS effectively suppressed ATP content in PDO tissues, with a fitted dose-response curve yielding an IC50 value of 2.49 µmol/L. In the RBE cell model, PQS inhibited RBE cell viability in a concentration-dependent manner, and the fitted dose-response curve yielded an IC50 value of 1.05 µmol/L. Furthermore, PQS at concentrations of 1, 2, and 4 µmol/L significantly suppressed colony formation of RBE cells, arrested the cell cycle at the S phase, induced apoptosis, and downregulated the expression of proteins associated with the NF-κB signaling pathway. Conclusion ICC PDO models were successfully established, confirming the anti-ICC activity of PQS. PQS inhibited RBE cell proliferation, potentially via the NF-κB signaling pathway, by arresting the cell cycle at the S phase and inducing apoptosis of RBE cells -
图 3 PQS对PDO生长的抑制活性(比例尺=500 µm,$ \bar{x} $±s,n=3)
A.不同PQS浓度处理的PDO形态变化;B.PQS不同浓度下PDO组织ATP含量;C.PQS抑制PDO生长浓度-响应拟合曲线**P<0.01,与细胞对照组比较。
图 4 PQS结构及其对RBE细胞存活率的影响($\bar x $±s, n=3)
A.PQS的化学结构;B.不同浓度PQS对细胞存活率的影响;C.不同浓度顺铂对细胞活存活率的影响
图 5 PQS对RBE细胞集落形成的影响($\bar x $±s, n=3)
A.细胞集落形成图;B.集落数定量分析结果 *P<0.05,**P<0.01,***P<0.001,与细胞对照组比较。
图 6 PQS对RBE细胞周期和细胞周期蛋白表达的影响($\bar x $±s, n=3)
A、B.流式细胞术检测细胞周期结果; C、D、E.Western印迹法检测CDK2和CDK4蛋白表达结果 *P<0.05,**P<0.01,与细胞对照组比较。
图 7 PQS对RBE细胞凋亡的影响($\bar x $±s, n=3)
A.流式细胞术检测细胞凋亡的散点图;B.凋亡细胞率统计结果***P<0.001,与细胞对照组比较。
图 8 PQS对RBE细胞中P65、P50蛋白表达的影响($\bar x $±s, n=3)
A.Western印迹法检测P65、P50蛋白条带图; B、C.分别为P65、P50蛋白表达半定量分析结果 *P<0.05,与细胞对照组比较。
表 1 所构建患者来源类器官(PDO)的患者临床信息
PDO编号 性别 年龄/岁 肿瘤类型 pT pN pM 分期 PDO#1 男 48 ICC T1 N0 M0 Ⅰ PDO#2 女 74 ICC T2 N1 M0 ⅢB ICC:肝内胆管癌;pT:病理T分期;pN:病理N分期;pM:病理M分期;T1:肿瘤直径≤2cm,无血管侵犯;T2:肿瘤直径>2cm,或伴有血管侵犯;N0:无区域淋巴结转移;N1:有区域淋巴结转移;M0:无远处转移;Ⅰ期 / ⅢB期根据TNM分期系统诊断。  下载: 导出CSV
下载: 导出CSV
-
[1] 陶雯琦, 刘颖斌. 晚期肝内胆管癌综合治疗研究进展[J]. 肿瘤, 2025, 45(3): 307-316. [2] SIEGEL R L, MILLER K D, FUCHS H E, et al. Cancer statistics, 2022[J]. CA A Cancer J Clin, 2022, 72(1): 7-33. doi: 10.4324/9781003281900-16 [3] 王政禄, 丛文铭. 肝移植常见肝脏恶性肿瘤的病理分类及其临床特点[J]. 实用器官移植电子杂志, 2019, 7(2): 122-127. [4] QURASHI M, VITHAYATHIL M, KHAN S A. Epidemiology of cholangiocarcinoma[J]. Eur J Surg Oncol, 2025, 51(2): 107064. doi: 10.1016/j.ejso.2023.107064 [5] BANALES J M, CARDINALE V, CARPINO G, et al. Cholangiocarcinoma: current knowledge and future perspectives consensus statement from the European Network for the Study of Cholangiocarcinoma(ENS-CCA)[J]. Nat Rev Gastroenterol Hepatol, 2016, 13(5): 261-280. doi: 10.1038/nrgastro.2016.51 [6] AKATEH C, EJAZ A M, PAWLIK T M, et al. Neoadjuvant treatment strategies for intrahepatic cholangiocarcinoma[J]. World J Hepatol, 2020, 12(10): 693-708. doi: 10.4254/wjh.v12.i10.693 [7] MORIS D, PALTA M, KIM C, et al. Advances in the treatment of intrahepatic cholangiocarcinoma: an overview of the current and future therapeutic landscape for clinicians[J]. CA A Cancer J Clin, 2023, 73(2): 198-222. doi: 10.3322/caac.21759 [8] CHANDRA P, SONG Y, DRILL E, et al. Intrahepatic cholangiocarcinoma: recurrence patterns, genomics, and survival[J]. JCO Precis Oncol, 2025, 9: e2500402. [9] GUJARATHI R, PESHIN S, ZHANG X C, et al. Intrahepatic cholangiocarcinoma: insights on molecular testing, targeted therapies, and future directions from a multidisciplinary panel[J]. Hepatol Commun, 2025, 9(7): e0743. doi: 10.1097/hc9.0000000000000743 [10] 董志涛, 朱妍静, 钱尤雯, 等. 肝内胆管细胞癌类器官对吉西他滨联合顺铂化疗方案疗效预测价值研究[J]. 中国实用外科杂志, 2025, 45(8): 931-937. doi: 10.19538/j.cjps.issn1005-2208.2025.08.15 [11] 林圣哲, 张灵玉, 陶世莉, 等. 胆管癌类器官模型的构建及药物反应异质性的初步分析[J]. 中国肿瘤生物治疗杂志, 2025, 32(11): 1121-1127. [12] DWYER B J, TIRNITZ-PARKER J E E. Patient-derived organoid models to decode liver pathophysiology[J]. Trends Endocrinol Metab, 2025, 36(3): 235-248. doi: 10.1016/j.tem.2024.07.019 [13] XU X, HUANG X F, XU W H. Marine actinomycetes-derived angucyclines and angucyclinones with biosynthesis and activity: past 10 years(2014–2023)[J]. Eur J Med Chem, 2025, 283: 117161. doi: 10.1016/j.ejmech.2024.117161 [14] 代影, 杨帆, 林厚文. 极地放线菌次级代谢产物及药理活性研究进展[J]. 天然产物研究与开发, 2023, 35(5): 901-913. doi: 10.16333/j.1001-6880.2023.5.018 [15] GIORDANO D, COPPOLA D, Russo R, et al. Marine microbial secondary metabolites: pathways, evolution and physiological roles[J]. Adv Microb Physiol, 2015, 66: 357-428. [16] VLACHOGIANNIS G, HEDAYAT S, VATSIOU A, et al. Patient-derived organoids model treatment response of metastatic gastrointestinal cancers[J]. Science, 2018, 359(6378): 920-926. doi: 10.1126/science.aao2774 [17] SELVARAJ B, WOON KIM D, PARK J S, et al. Neuroprotective effects of 2-heptyl-3-hydroxy-4-quinolone in HT22 mouse hippocampal neuronal cells[J]. Bioorg Med Chem Lett, 2021, 49: 128312. doi: 10.1016/j.bmcl.2021.128312 [18] NANDAKUMAR N, DANDELA R, GOPAS J, et al. Quorum sensing modulators exhibit cytotoxicity in Hodgkin’s lymphoma cells and interfere with NF-κB signaling[J]. Bioorg Med Chem Lett, 2017, 27(13): 2967-2973. doi: 10.1016/j.bmcl.2017.05.012 [19] JIA T Y, LI F M, LI T Z, et al. A Pseudomonas aeruginosa quorum-sensing metabolite manipulates macrophage ferroptosis through a methylation pathway[J]. Nat Commun, 2025, 16: 9992. doi: 10.1038/s41467-025-65142-y [20] OGBECHI J, HUANG Y S, CLANCHY F I L, et al. Modulation of immune cell function, IDO expression and kynurenine production by the quorum sensor 2-heptyl-3-hydroxy-4-quinolone(PQS)[J]. Front Immunol, 2022, 13: 1001956. doi: 10.3389/fimmu.2022.1001956 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 900
- HTML全文浏览量: 465
- PDF下载量: 7
- 被引次数: 0
