

 下载:
下载:

-
阿尔茨海默病(Alzheimer’s disease, AD)是一种以进行性认知功能衰退、精神行为异常及日常生活能力下降为主要特征的神经退行性疾病。在当前全球老龄化程度日益加深的背景下,AD的发病率呈现出持续上升的态势[1]。据预测,至2050年全球AD患病人数将达1.39亿[2]。我国AD患者约占全球总数的25.5%,伴随人口老龄化进程的深化,其发病率、致残率及死亡率逐年攀升,预计2050年患病人数将突破
2765 万[3]。目前,美国食品药品监督管理局批准用于治疗AD的药物主要包括乙酰胆碱酯酶抑制剂(AChEIs)、N-甲基-D-天门冬氨酸拮抗剂[4],以及最新上市的仑卡奈单抗、阿杜卡单抗等药物[5-6]。基于“多组分、多途径、多靶点”的协同作用特点,中药在AD临床干预中展现出独特优势[7]。然而,中药的复杂成分也为系统阐释其治疗机制带来挑战,需要科学的研究方法解析中药治疗AD的机制。目前用于阐释中药治疗AD的分子机制的方法,包括表面等离子共振、生物膜干涉等分子互作实验技术以及代谢组学、网络药理学等系统生物学分析方法。其中,细胞代谢组学通过在微观层面研究细胞在不同生理状态下代谢物种类、含量、代谢通路等方面的动态变化,揭示细胞代谢网络的复杂调控机制,进而揭示中药干预AD的疗效及作用原理,为深入理解中药治疗AD的分子机制提供重要依据[8]。本文综述治疗AD的现有药物及中药干预AD的研究进展,为AD发病机制的研究和治疗药物的开发提供科学依据。 -
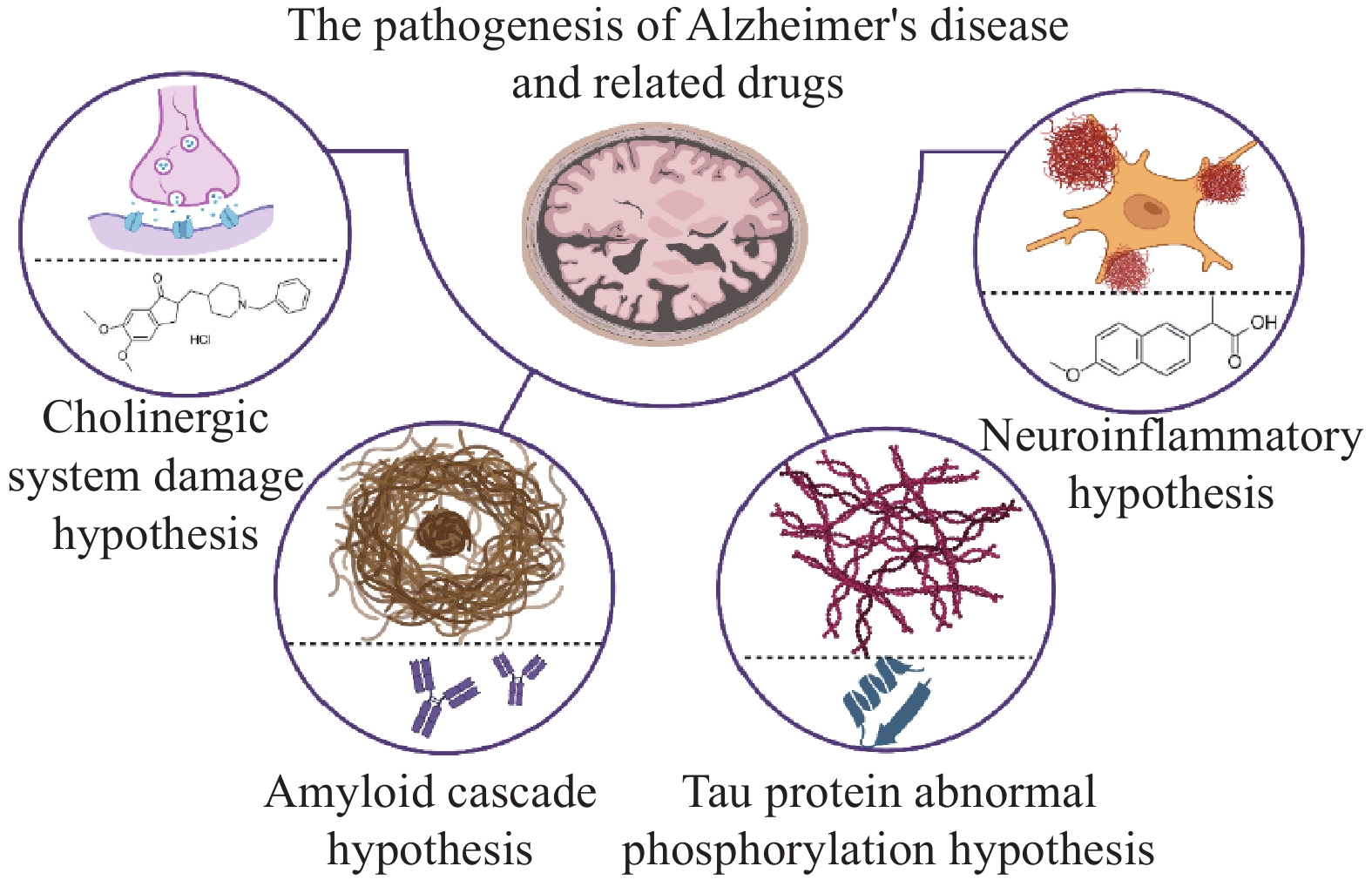
AD的发病机制复杂,当前主流假说有中枢胆碱能系统损伤、淀粉样蛋白(Aβ)级联反应、Tau蛋白异常磷酸化、炎症损伤等假说。尽管这些理论从不同生物学维度阐释了疾病发生发展机制,但尚未形成能够全面覆盖疾病病理特征的统一机制模型。基于上述假说的药物研发,是AD治疗领域的重要探索方向,如图1所示。然而现阶段的AD治疗药物,仅能在一定程度上改善症状,且伴随一定不良反应,对逆转病理进程及改善疾病转归作用有限。
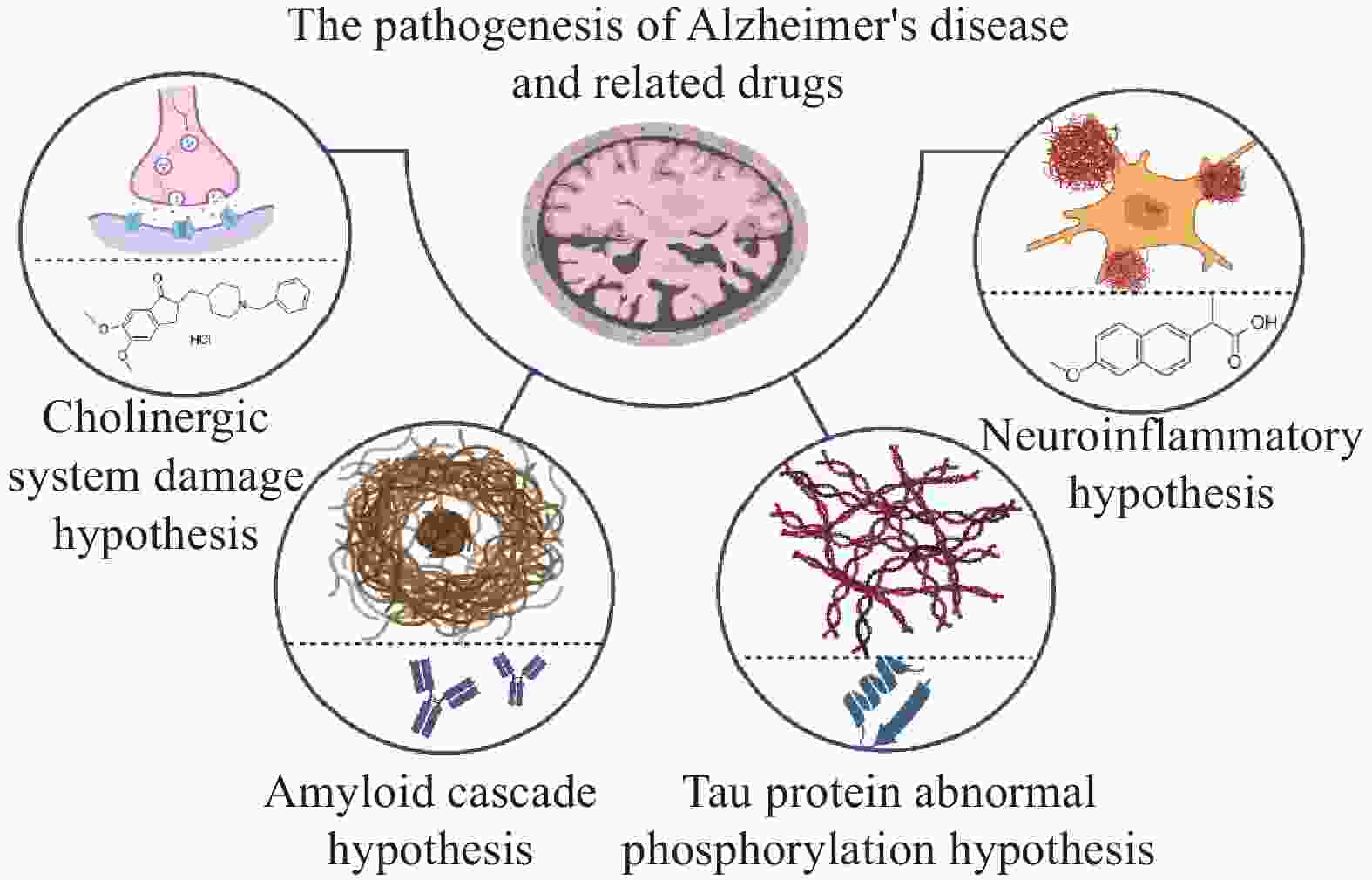
图 1 阿尔茨海默病的发病机制假说及相关药物
-
中枢胆碱能系统损伤假说是最早被提出用于阐释AD发病机制的理论。该假说指出,麦纳尔基底核中胆碱能神经元发生显著退行性病变,导致其主要投射区域——大脑皮层和海马区的胆碱乙酰转移酶(ChAT)活性显著降低,致使患者突触间隙的乙酰胆碱(ACh)水平下降[9]。ACh含量不足与AD患者的学习记忆能力减退及认知功能衰退存在密切关联。AChEIs是临床干预AD的重要药物类别,其作用机制为通过抑制AChE活性维持ACh的有效浓度,改善神经传递功能。目前临床常用的AChEIs包括多奈哌齐、卡巴拉汀及加兰他敏等,其中多奈哌齐因患者耐受性更佳、疗效更稳定,成为当前AD治疗中应用最广泛的药物之一。然而,AChEIs的治疗效果高度依赖AD患者脑内胆碱能神经元的结构与功能完整性,随着AD病情进展至重度阶段,胆碱能神经元已出现广泛变性坏死,因此此类药物不适用于重度AD患者。此外,在针对轻中度AD患者的长期治疗中,此类药物的有效率仅约50%,且停药后疗效维持时间仅为1~1.5年[10]。中枢胆碱能系统损伤假说从机制上解释了AD的发病与ACh水平下降的关联,基于此研发的AChEIs成为AD临床治疗的重要药物,但这类药物存在适用阶段有限、长期疗效欠佳、无法逆转病理等局限。
-
淀粉样蛋白级联假说是AD发病机制的核心理论之一,该假说认为淀粉样前体蛋白(Amyloid Precursor Protein,APP)经过β分泌酶和γ分泌酶连续酶切后,生成具有神经毒性的Aβ片段。在AD病理进程中,Aβ的产生与清除平衡被打破,导致Aβ在脑实质内异常聚集,形成淀粉样斑块,并通过氧化应激、神经炎症等机制引发神经元功能紊乱[11]。基于该假说,抗Aβ单克隆抗体——阿杜卡单抗和仑卡奈单抗先后获得FDA的批准,用于AD源性轻度认知障碍和轻度AD的治疗。有研究对抗体药物与Aβ单体、Aβ纤维、Aβ原纤维的结合能力进行研究,发现阿杜卡单抗和仑卡奈单抗与Aβ单体亲和力均较低,仑卡奈单抗与Aβ原纤维的结合能力比与纤维的结合能力强10倍;而阿杜卡单抗更倾向于结合Aβ纤维,这解释了这些抗体在疗效和副作用方面的临床结果差异[12]。目前,单抗类药物的随机对照试验仅在轻度或早期AD患者中进行,缺乏不同病情分期的样本量。同时,单抗类药物普遍采用注射给药,其便携性及依从性存在明显局限。此外,以Aβ为靶点的小分子药物研发也取得突破,以贝沙罗汀为先导化合物研发出的OAB-14干混悬剂,对脑内Aβ的代谢清除有明显促进作用[13]。
-
微管相关蛋白Tau,又简称Tau蛋白,其异常磷酸化假说是AD病理机制的重要理论假说。在正常生理状态下,Tau蛋白与磷酸化微管相关蛋白处于相对平衡状态,维持细胞结构稳定性。AD病理进程中,Tau蛋白发生异常磷酸化,导致其与微管分离并在神经元内蓄积,形成神经原纤维缠结[14]。目前,针对Tau蛋白异常磷酸化的治疗策略研究众多,如Tau蛋白激酶抑制剂GSK-3β、Tau蛋白沉积抑制剂LMTX及Tau蛋白抗体JNJ-
63733657 、主动免疫疫苗AADvac1等[15-16]。其中AADvac1疫苗是一种靶向Tau蛋白的活性肽疫苗,可激活免疫系统产生抗Tau抗体,从而减少神经纤维缠结的形成,延缓或阻止疾病的进展。该疫苗在降低神经退行性标志物方面表现出潜力,但AADvac1对认知功能改善效果未达统计学显著性[17]。多种中药复方及活性单体化合物被发现能够有效逆转失衡的Tau蛋白水平,如人参皂苷Rbl能降低Tau蛋白的磷酸化水平,促进Tau蛋白去磷酸化增多而稳定神经元的微管系统,进而有可能逆转AD的病理改变,对AD的治疗达到一定效果[18]。 -
神经炎症假说是AD病理机制的重要补充理论。该假说认为,AD患者脑内Aβ沉积及神经纤维缠结可与小胶质细胞表面受体结合,诱导小胶质细胞过度激活,导致促炎因子大量释放;这些促炎因子反过来加剧脑中Aβ、Tau蛋白积累,形成“病理-炎症”恶性循环,推动AD病程进展[19]。针对该假说,PPARγ激动剂如吡格列酮、选择性COX-2抑制剂如塞来昔布、非选择性COX抑制剂吲哚美辛等被用来干预神经炎症,在AD治疗中展现出一定的获益。但由于临床获益证据不足且存在胃肠道损伤、心血管风险等安全性问题,其用于AD治疗药物的有效性和安全性仍存争议[20]。研究发现连翘酯苷B能够调节脑内神经炎症等相关信号通路,缓解由AD造成的认知障碍。在APP/PS1双转基因小鼠模型中,连翘酯苷B给药能显著改善小鼠的空间学习记忆能力及认知能力,同时伴随脑内Aβ斑块面积缩小、Tau蛋白磷酸化水平降低,以及小胶质细胞和星形胶质细胞活化程度的明显抑制[21]。
-
化学药和生物药治疗多针对单一靶点,无法覆盖AD多因素致病、多环节进展的病理本质。同时治疗过程伴随多种不良反应,如化学药的胆碱酯酶抑制剂易造成胃肠反应、胆碱酯酶抑制剂只适用于肝肾功能良好、无心脏疾病的轻中度AD患者。而中药以其独特的多成分、多靶点作用特点为AD防治研究提供了重要方向。目前,诸多中药复方、单味药材、以及药材的单体化合物在AD治疗研究中已取得显著进展,为疾病的多元化干预提供了临床依据。
有研究采用代谢组学技术研究了定志小丸防治AD的作用和机制,在AD模型大鼠中鉴定出26个内源性生物标志物,发现定志小丸主要调控牛磺酸和亚牛磺酸代谢、色氨酸代谢和苯丙氨酸代谢的通路,可能通过介导抗氧化应激、调节能量代谢、改善肠道微生物及保护神经细胞来发挥治疗AD的作用[22]。本课题组采用一种整合GEO数据集、网络药理学及多平台代谢组学的系统性研究策略,揭示经典名方不忘散通过调控氨基糖和核苷酸糖代谢、甘油磷脂代谢等通路,并作用于ARG1、MAOA、AChE、XDH、GAD2等关键靶点发挥AD治疗作用[23]。此外,人参与石菖蒲的药对配伍可促进活性成分透过血脑屏障,通过抗炎、抗氧化应激及调节神经元-突触可塑性等多途径延缓AD进程,体现了中药复方多层次、多系统、多靶点的作用特点[24]。关于单味药材研究,网络药理学、代谢组学等系统生物学方法为阐释其作用机制提供了新视角。有研究通过基于质谱成像技术的空间代谢组学方法全面分析人参治疗AD的机制,发现人参有缓解Aβ蛋白沉积和tau蛋白磷酸化、增强突触再生能力以及抑制炎症反应的作用,研究发现AD大鼠中共有19种生物标志物被显著回调,涉及多种氨基酸代谢途径[25]。多项研究表明,AD存在显著的神经元铁凋亡,抑制神经元铁凋亡对认知表现和类AD样病理有显著影响[26]。有研究通过整合网络药理学和多组学技术,揭示了石菖蒲通过激活Nrf2/GPX4轴来减少铁沉积并抑制海马神经铁块状病变的药理机制,提升了细胞存活率,减少了细胞内游离铁、活性氧和脂质过氧化,最终在体外和体内水平提升了APP/PS1小鼠的认知表现[27]。
大量研究证实,中药中不同活性成分可通过针对特异靶点发挥防治AD的药效。人参中多种人参皂苷可调节Tau蛋白磷酸化平衡改善突触可塑性[28]。人参衍生的糖脂蛋白含有G蛋白耦合的溶血酸,具有减少类转β淀粉样蛋白沉积、减轻血脑屏障的功能损伤等[29-30]。石菖蒲中的β-细辛醚显著提高了海马区超氧化物歧化酶和谷胱甘肽过氧化物酶的活性,减轻了AD的神经细胞丧失,显示出其在减轻氧化应激和神经损伤方面的保护作用[31]。对于茯苓中有效成分茯苓酸,有研究通过HE染色、免疫组化等实验技术,证明茯苓酸可改善AD大鼠神经炎症,其作用机制与抑制HMGB1/TLR4通路相关[32]。
尽管中药在AD治疗中展现出良好疗效,但其具体作用机制亟待深入研究。传统亲和力检测技术存在预设靶点、难以反映真实生理效应等局限,而组学等生物信息学手段可实现AD细胞及亚细胞水平代谢紊乱的精准解析,为揭示中药调控AD细胞代谢的时空特异性机制与创新药研发提供支撑。以细胞代谢组学为代表,通过捕捉中药干预后AD模型细胞内代谢物的种类、含量及通路变化,揭示中药调控细胞代谢网络的作用机制,为系统阐明中药活性成分的作用靶点、信号通路及代谢网络提供了有力工具,有助于阐明中药作用于AD的疗效及机制,推动中药在AD治疗领域的应用和发展。
-
细胞代谢组学作为代谢组学的分支领域,主要考察细胞代谢变化规律,揭示健康和病理生物体之间细胞异质性及生化差异[33]。基于AD细胞模型的细胞代谢组学研究为生物体生理和病理研究提供更准确、更丰富的信息,有效补充整体代谢组学的不足[34]。
-
AD细胞模型通过模拟Aβ沉积、Tau蛋白异常磷酸化、神经炎症等核心病理特征,为AD机制解析和药物筛选提供了可控、可重复的实验平台,主要包括干细胞模型、神经细胞系模型及脑类器官模型等。随着干细胞技术、微流控芯片及类器官技术的发展,AD细胞模型已从传统的二维神经细胞系模型,向更接近人体病理的三维类器官模型进阶,在AD机制探索及药效评价中的作用愈发凸显。
-
干细胞模型包括诱导多能干细胞模型、间充质干细胞外泌体模型以及胚胎干细胞模型等。这类模型凭借强大的分化潜能、良好的细胞均一性,以及可匹配患者特异性遗传背景的独特优势,已成为大规模药物筛选和疾病机制研究的重要工具[35]。有研究将诱导多能干细胞分化为人类神经元并移植至人源化APOE3或APOE4敲入小鼠的海马区后,发现APOE基因型显著影响Aβ沉积和Tau蛋白病理的严重程度;进一步的单细胞RNA测序分析发现,诱导多能干细胞神经元分泌的APOE,尤其是APOE4亚型,在AD发病中发挥重要作用[36]。此外,多项研究表明干细胞及干细胞来源的细胞外囊泡在AD治疗中潜力显著[37-38]。间质干细胞可通过分化补充神经元、调节Aβ沉积与Tau磷酸化、抑制神经炎症等发挥作用,其EVs能减少Aβ斑块、改善认知[39]。通过冈田酸诱导SH-SY5Y细胞进行AD细胞造模,发现人嗅黏膜间充质干细胞共培养对于冈田酸诱导损伤SH-SY5Y细胞建立的AD模型细胞有治疗作用[40]。间质干细胞等干细胞模型因伦理限制较少成为研究热点,但仍面临致瘤性、移植效率等挑战,未来需进一步优化技术以推动临床转化。
-
神经细胞系模型主要基于PC12细胞、SH-SY5Y细胞和N2a细胞等,通过基因导入、药物诱导等建立AD模型。PC12细胞是大鼠肾上腺髓质嗜铬细胞瘤细胞系,对神经生长因子有可逆的神经元显形反应,通过处理PC12细胞可以诱导其出现类似于AD造成的细胞损伤和死亡现象,常用于研究AD的分子机制和药物筛选[41]。SH-SY5Y是人类神经母细胞瘤细胞系,N2a是小鼠神经瘤细胞系,两者均适合基因修饰实验,通过诱导这些细胞产生Aβ或Tau蛋白异常,可模拟AD的病理特征,广泛应用于研究神经毒性机制和药物干预效果评价[42]。有研究通过视黄酸和脑源性神经营养因子诱导SH-SY5Y细胞建立AD体外模型,研究发现3种淀粉样β肽(KLVFF、RGKLVFFGR和RIIGL)对该模型的不同作用,发现RIIGL肽对正常SH-SY5Y细胞的细胞毒性活性最低,而对AD模型细胞则表现出较高的细胞毒性,并证明RIIGL肽通过调节淀粉样β肽形成、神经元凋亡、神经酰胺分解和氧化磷酸化作用于分化的SH-SY5Y,这说明RIIGL肽有用于治疗AD的潜力[43]。
-
脑类器官模型通过干细胞的自组织能力形成三维结构,精准模拟AD病理微环境,接近真实的脑组织,为AD研究提供了高度仿生的实验平台。在AD机制研究中,该模型能够精准再现Aβ斑块沉积、神经原纤维缠结形成等核心病理特征,为解析AD病理进程提供了直观的体外模型。此外,通过结合微流控芯片技术,脑类器官模型可实现对不同脑区在疾病进展中的交互作用及药物对脑组织的综合影响的动态研究[44]。利用人类诱导多能干细胞衍生的脑类器官模拟血脑屏障破坏后的环境,研究其在散发性AD中的应用;通过优化人类诱导多能干细胞诱导和脑类器官培养流程,生成具有皮质结构的脑类器官,再将其暴露于血清中,成功模拟了散发性AD的病理特征,该模型为研究散发性AD的发病机制及药物综合效应提供了新平台[45]。为研究司美格鲁肽在AD中的治疗效果和机制,基于小鼠和人类脑类器官模型,发现司美格鲁肽能够显著减少Tau蛋白的磷酸化和Aβ的积累,进而发挥AD治疗效果[46]。
AD细胞模型作为AD研究的关键支撑平台,涵盖神经细胞系模型、干细胞模型及脑类器官模型三类核心体系,可模拟AD核心病理特征。未来,AD细胞模型的构建应不局限于单一细胞的培养,而是迈向多细胞系统协同,例如可通过CRISPR基因编辑[47],以精准的基因组编辑复刻散发性AD的多基因位点变异,提升AD细胞模型对临床病例的代表性。
-
细胞代谢组学作为聚焦细胞层面的研究手段,通过精准解析细胞内源性代谢物动态变化,揭示生理与病理状态下的生化差异及细胞异质性。相较于传统体液代谢组学,细胞代谢组学在AD研究中具备时空分辨率高、可定位神经元等特定脑细胞的代谢紊乱、直接关联代谢物变化等研究优势,与AD核心病理紧密相关,已逐渐应用于AD发病机制、代谢重编程、药物靶点的发现等研究。
脂质代谢组学作为细胞代谢组学的重要分支,聚焦细胞内脂质类代谢物的组成、含量及通路调控以解析脂质代谢紊乱相关机制。有研究以过表达APP的瑞典家族突变的N2a细胞作为AD的体外模型,通过高分辨液相色谱-质谱技术与气相色谱-质谱联用分析其脂质谱差异,描绘线粒体相关膜和线粒体的脂质代谢图谱。发现在AD细胞中,线粒体相关膜等组分的脂质稳态发生特异性改变,而这些脂质组学的变化与线粒体功能障碍、氧化应激等AD病理特征相关联,揭示了疾病状态下亚细胞层面的代谢异常[48]。Aβ在AD中的聚集特征表现为线粒体功能障碍和线粒体自噬受损。有研究者基于Aβ水平的线粒体自噬与ATP相关细胞代谢组学,发现Aβ积累可抑制线粒体自噬,导致糖酵解、三羧酸循环的中间产物异常生成及代谢紊乱,揭示线粒体功能异常与Aβ的关联,为AD能量代谢障碍机制提供依据[49]。研究者基于AD患者成纤维细胞转化的功能性诱导神经元开展研究,利用质谱分析发现功能性诱导神经元的细胞质基质中,糖酵解中间产物大量积累,乳酸水平显著升高,该结果与转录组选择性剪接分析共同提示AD神经元出现类似肿瘤的Warburg效应,揭示了PKM2作为核心靶点驱动的代谢重编程及表观遗传调控在AD神经元退行中的核心作用[50]。为探究丹参防治AD的药效机制,本课题组采用40 nmol/L冈田酸诱导SH-SY5Y细胞引起Tau 蛋白异常磷酸化建立AD细胞模型,并运用细胞代谢组学寻找AD细胞模型与SH-SY5Y细胞的差异代谢物,基于UHPLC-QTOF/MS发现在丹参作用下,有30个代谢物水平有向正常组回调的趋势,其中18个代谢物与模型组比较有显著差异,包括色氨酸、苯丙氨酸、乳酸、柠檬酸、丙酮酸等,证明丹参对神经递质生成及免疫激活的调节有显著作用[51]。
-
近年来,单细胞研究、质谱成像等前沿技术的突破,为细胞代谢组学的精准化发展提供了关键支撑,其中单细胞代谢组学实现了对细胞代谢过程的单细胞水平解析。传统代谢组学以群体细胞为研究对象,所获取的代谢数据实质是细胞群的均值化表征,故而难以规避细胞间异质性信息的遮蔽与丢失。而单细胞代谢组学技术可精准捕获单个细胞的代谢特征,不仅能够有效揭示细胞群体内的代谢异质性与表型多样性,更能提供相比基因组学与转录组学更丰富的细胞功能表型信息。该技术主要由单细胞分离与捕获、单细胞代谢物分析及代谢数据处理三个核心环节构成,其显著优势在于有效地避免由于细胞异质性引起的误差,可反映单一细胞功能以及揭示细胞异质性与代谢间的关联。
AD存在显著的细胞代谢异质性,不同细胞类型的代谢紊乱特征存在明显差异:神经元表现为能量代谢与脂质代谢相关活动的显著下调,而微胶质细胞则呈现出与神经炎症相关的代谢活性增强[52]。研究通过整合单细胞技术与相互作用组学方法,成功筛选出数百个AD相关细胞类型特异性主代谢调节因子,其中神经元与微胶质细胞间的代谢调控相互作用最为突出,直接证实了AD细胞代谢异质性研究对阐明疾病机制的关键意义[53]。APOE4是AD的关键遗传风险因素,可通过激活胞质磷脂酶A2等通路干扰脑内脂质代谢、维持慢性炎症,通过单细胞类型脂质组学解析脑部炎症和脂质代谢的关系,提出多种脑细胞类型,如星形胶质细胞、小胶质细胞和神经元,连同神经血管单元,发展出具有不同特征的炎症表型[54]。为实现单细胞水平脂质谱的高分辨解析,研究人员发展了荧光引导序贯单细胞MALDI-ICC质谱技术,对逾千个单细胞开展脂质组学分析与分子注释。结果显示,磷脂酰胆碱类脂质在少突胶质细胞与神经元中显著富集,其相对丰度明显高于星形胶质细胞;六糖神经酰胺在这三类细胞中表达均有差异;突触前神经元富集磷脂酰乙醇胺,非突触前神经元脂质组成更丰富[55]。这些具有细胞类型特异性的脂质组学“指纹”,不仅直接影响细胞功能状态,为AD靶向干预措施的开发提供了潜在分子靶点。单细胞代谢组学通过解析细胞异质性与代谢、脂质特征的关联,为AD的病理机制研究提供了精准视角。
质谱成像(MSI)凭借高灵敏度和高分辨率的特点,可通过分析AD患者或者模型生物样本中生物分子的空间分布筛选潜在生物标志物,并可视化代谢物与脂质的空间化学变化,为揭示AD的代谢异常、脂质稳态失衡等病理机制及诊断提供科学依据,进一步拓展了AD研究的空间维度[56]。目前,基于单细胞MALDI质谱成像技术广受关注,但缺乏针对大细胞群体中 < 200Da低质量亲水性代谢物的细胞代谢组学分析方法。研究建立了一种小分子成像重扫质谱的技术,通过质量引导的MSI工作流程,实现对氨基酸等低质量代谢物的MSI分析和细胞水平MS2识别;并发现器官型海马切片培养的内源性微胶质细胞中,LPS激活涉及乙烯酸与牛磺酸比率变化及神经元与胶质细胞间谷氨酰胺-谷氨酸转运改变[57]。
综上,单细胞代谢组学与质谱成像技术的迭代突破,为细胞代谢组学的精准化发展奠定核心支撑,更开辟了AD病理机制研究与精准诊断的全新视角。随着技术的不断进步,细胞代谢组学方法也在不断演进,新一代的质谱和核磁共振技术的应用,以及大数据分析和AI的发展,不断提高细胞代谢组的精密度与准确度,为AD药物研究提供更多选择。
-
AD发病率随全球老龄化持续攀升,我国患病人数占比高且增长趋势严峻,而现有药物仅能改善症状或延缓进展,无法逆转病理进程,因此研发新型治疗药物迫在眉睫。中药凭借“多组分、多途径、多靶点”的独特优势,在AD防治中展现出显著潜力。细胞代谢组学作为聚焦微观层面的核心技术,通过AD细胞模型,干细胞模型、神经细胞系模型、脑类器官模型的进阶应用,结合单细胞代谢组学、质谱成像等前沿手段,有效弥补了整体代谢组学的不足,揭示AD病理中脂质代谢异常、线粒体功能障碍致能量代谢紊乱、代谢重编程等关键机制,实现对细胞异质性、代谢物空间分布的精准解析,为AD病理机制研究与药物靶点发现提供了全新视角。然而,细胞代谢组学存在单细胞捕获效率低、低丰度代谢物检测灵敏度不足、代谢物成像技术不完善等问题。未来通过研发高效单细胞捕获与高灵敏度检测技术,完善代谢物成像方法,整合细胞代谢组学与多技术的协同应用,有望推动AD机制研究向精准化、系统化发展,为开发安全有效的AD治疗策略提供科学支撑。
Progress in drug research for Alzheimer’s disease and application of cellular metabolomics
-
摘要: 阿尔茨海默病(Alzheimer’s disease,AD)是一种典型的神经退行性疾病,致病机理尚未完全明确。目前临床治疗AD的药物有限,且不良反应较多。中药具有多成分、多靶点的特点,对于AD防治有独特的优势和潜力。随着对中药治疗AD优势的深入认识,探索其作用机制成为推动中药现代化应用的关键环节。目前研究技术有分子生物学、网络药理学、质谱成像与细胞代谢组学等。细胞代谢组学通过研究细胞内代谢物变化,精准捕捉细胞在不同生理状态下代谢物种类、含量及代谢通路等方面的动态变化,有望揭示细胞内中药代谢网络的复杂调控机制。基于AD细胞模型的代谢组学研究有助于发现AD特异性标志物,进一步探究AD发病和防治机制。本文对AD防治药物,以及近年来细胞代谢组学在AD研究中的应用进展进行综述,以期为AD机制研究、临床诊治和药物研发提供参考依据。Abstract: Alzheimer’s disease (AD) is a typical neurodegenerative disorder, and its pathogenic mechanism remains to be fully elucidated. Currently, there are limited drugs available for the clinical treatment of AD, and many of these drugs are associated with numerous adverse effects. Traditional Chinese medicine (TCM), featuring multi-component and multi-target properties, possesses unique advantages and potential in the prevention and treatment of AD. AD cell models enable in-depth investigations into the pathogenesis of AD and the mechanisms underlying drug-based prevention and treatment. Cell metabolomics studies changes in intracellular metabolites to accurately capture dynamic variations in metabolite types, contents, and metabolic pathways of cells under different physiological states; this approach holds promise for revealing the complex regulatory mechanisms of cellular metabolic networks, identifying AD-specific biomarkers, and providing support for research on the mechanisms of AD. This article reviews the drugs used for AD prevention and treatment, as well as recent progress in the application of cellular metabolomics in AD research, aiming to provide a reference for studies on AD mechanisms, clinical diagnosis and treatment, and drug development.
-
[1] AARSLAND D, SUNDE A L, TOVAR-RIOS D A, et al. Prevalence of Alzheimer’s disease’s pathology in the community[J]. Nature, 2025: 1-5. [2] 徐勇, 王军, 王虹峥, 等. 2023中国阿尔茨海默病数据与防控策略[J]. 阿尔茨海默病及相关病, 2023, 6(3): 175-192. [3] VAN GIAU V, BAGYINSZKY E, YANG Y S, et al. Genetic analyses of early-onset Alzheimer’s disease using next generation sequencing[J]. Sci Rep, 2019, 9(1): 8368. doi: 10.1038/s41598-019-44848-2 [4] CONTI FILHO C E, LOSS L B, MARCOLONGO-PEREIRA C, et al. Advances in Alzheimer’s disease’s pharmacological treatment[J]. Front Pharmacol, 2023, 14: 1101452. doi: 10.3389/fphar.2023.1101452 [5] BUDD HAEBERLEIN S, AISEN P S, BARKHOF F, et al. Two randomized phase 3 studies of aducanumab in early Alzheimer’s disease[J]. J Prev Alzheimers Dis, 2022, 9(2): 197-210. doi: 10.14283/jpad.2022.30 [6] VAN DYCK C H, SWANSON C J, AISEN P, et al. Lecanemab in early Alzheimer’s disease’s[J]. N. Engl. J. Med., 2023, 388(1): 9-21. doi: 10.1056/NEJMoa2212948 [7] CHEN S Y, GAO Y, SUN J Y, et al. Traditional Chinese medicine: role in reducing β-amyloid, apoptosis, autophagy, neuroinflammation, oxidative stress, and mitochondrial dysfunction of Alzheimer’s disease[J]. Front Pharmacol, 2020, 11: 497. doi: 10.3389/fphar.2020.00497 [8] 戴建英, 王辉, 刘敏, 等. 阿尔茨海默病的病理模型及代谢组学研究进展[J]. 药学服务与研究, 2021, 21(4): 241-247,320. [9] CHEN Z R, HUANG J B, YANG S L, et al. Role of cholinergic signaling in Alzheimer’s disease[J]. Molecules, 2022, 27(6): 1816. doi: 10.3390/molecules27061816 [10] 金艳, 李阳, 雷旭伟. 多奈哌齐治疗AD源性轻度认知功能障碍患者的临床研究[J]. 中国临床药理学杂志, 2024, 40(18): 2616-2620. doi: 10.13699/j.cnki.1001-6821.2024.18.002 [11] YEAPURI P, MACHHI J, LU Y M, et al. Amyloid-β specific regulatory T cells attenuate Alzheimer’s disease pathobiology in APP/PS1 mice[J]. Mol Neurodegener, 2023, 18(1): 97. doi: 10.1186/s13024-023-00692-7 [12] SÖDERBERG L, JOHANNESSON M, NYGREN P, et al. Lecanemab, aducanumab, and gantenerumab: binding profiles to different forms of amyloid-beta might explain efficacy and side effects in clinical trials for Alzheimer’s disease[J]. Neurotherapeutics, 2023, 20(1): 195-206. doi: 10.1007/s13311-022-01308-6 [13] ZHANG X, CAO R L, ZHU C, et al. Mechanism of anti-AD action of OAB-14 by enhancing the function of glymphatic system[J]. Neurochem Int, 2023, 171: 105633. doi: 10.1016/j.neuint.2023.105633 [14] MURALIDAR S, AMBI S V, SEKARAN S, et al. Role of tau protein in Alzheimer’s disease: The prime pathological player[J]. Int J Biol Macromol, 2020, 163: 1599-1617. doi: 10.1016/j.ijbiomac.2020.07.327 [15] GAUTHIER S, FELDMAN H H, SCHNEIDER L S, et al. Efficacy and safety of tau-aggregation inhibitor therapy in patients with mild or moderate Alzheimer’s disease: a randomised, controlled, double-blind, parallel-arm, phase 3 trial[J]. Lancet, 2016, 388(10062): 2873-2884. doi: 10.1016/S0140-6736(16)31275-2 [16] WISCHIK C M, STAFF R T, WISCHIK D J, et al. Tau aggregation inhibitor therapy: an exploratory phase 2 study in mild or moderate Alzheimer’s disease[J]. J Alzheimers Dis, 2015, 44(2): 705-720. doi: 10.3233/jad-142874 [17] NOVAK P, SCHMIDT R, KONTSEKOVA E, et al. Safety and immunogenicity of the tau vaccine AADvac1 in patients with Alzheimer’s disease: a randomised, double-blind, placebo-controlled, phase 1 trial[J]. Lancet Neurol, 2017, 16(2): 123-134. doi: 10.1016/S1474-4422(16)30331-3 [18] 杨淑达, 于浩飞, 张兰春, 等. 人参皂苷Rb1对Aβ1-42导致的Tau蛋白异常磷酸化的影响[J]. 天然产物研究与开发, 2020(7): 1143-1147,1098. doi: 10.16333/j.1001-6880.2020.7.008 [19] ROVETA F, BONINO L, PIELLA E M, et al. Neuroinflammatory biomarkers in Alzheimer’s disease: from pathophysiology to clinical implications[J]. Int J Mol Sci, 2024, 25(22): 11941. doi: 10.3390/ijms252211941 [20] DHAPOLA R, HOTA S S, SARMA P, et al. Recent advances in molecular pathways and therapeutic implications targeting neuroinflammation for Alzheimer’s disease[J]. Inflammopharmacology, 2021, 29(6): 1669-1681. doi: 10.1007/s10787-021-00889-6 [21] KONG F G, JIANG X, WANG R C, et al. Forsythoside B attenuates memory impairment and neuroinflammation via inhibition on NF-κB signaling in Alzheimer’s disease[J]. J Neuroinflammation, 2020, 17(1): 305. doi: 10.1186/s12974-020-01967-2 [22] HE Y, WANG Y M, LIU S, et al. A metabolomic study of the urine of rats with Alzheimer’s disease and the efficacy of Ding-Zhi-Xiao-Wan on the afflicted rats[J]. J Sep Sci, 2020, 43(8): 1458-1465. doi: 10.1002/jssc.201900944 [23] WANG H, CHAO L, SHEN S Q, et al. Exploring the pharmacological mechanism of Bu-Wang San on Alzheimer’s disease through multiple GEO datasets of the human hippocampus, network pharmacology, and metabolomics based on GC-MS and UPLC-Q/TOF-MS[J]. J Ethnopharmacol, 2025, 350: 119994. doi: 10.1016/j.jep.2025.119994 [24] 管秀菊, 侯翰如, 张立娟, 等. 人参-石菖蒲药对防治阿尔茨海默病的作用机制及临床应用[J]. 中医药导报, 2020, 26(12): 161-163,190. doi: 10.13862/j.cnki.cn43-1446/r.2020.12.040 [25] 范宇婷, 刘志强, 邢俊鹏, 等. 基于空间代谢组学方法研究人参治疗阿尔兹海默症大鼠的药效物质及作用机制[J]. 质谱学报, 2024, 45(6): 723-738. [26] STOCKWELL B R, FRIEDMANN ANGELI J P, BAYIR H, et al. Ferroptosis: a regulated cell death nexus linking metabolism, redox biology, and disease[J]. Cell, 2017, 171(2): 273-285. doi: 10.1016/j.cell.2017.09.021 [27] XIONG R, LIU H X, ZHANG S P, et al. Acorus tatarinowii Schott attenuates Alzheimer’s disease via neuronal ferroptosis inhibition: a synergistic network pharmacology and multi-omics profiling study[J]. J Ethnopharmacol, 2026, 356: 120829. doi: 10.1016/j.jep.2025.120829 [28] ZHAO C M, YUE J, XIE Y, et al. A ginsenoside composition ameliorated aβ and tau aggregation via autophagy lysosome pathway[J]. Mol Neurobiol, 2025, 62(9): 11822-11833. doi: 10.1007/s12035-025-05017-x [29] JAKARIA M, AZAM S, GO E A, et al. Biological evidence of gintonin efficacy in memory disorders[J]. Pharmacol Res, 2021, 163: 105221. doi: 10.1016/j.phrs.2020.105221 [30] JANG M, CHOI S H, CHOI J H, et al. Ginseng gintonin attenuates the disruptions of brain microvascular permeability and microvascular endothelium junctional proteins in an APPswe/PSEN-1 double-transgenic mouse model of Αlzheimer’s disease[J]. Exp Ther Med, 2021, 21(4): 310. doi: 10.3892/etm.2021.9741 [31] SAKI G, EIDI A, MORTAZAVI P, et al. Effect of β-asarone in normal and β-amyloid-induced Alzheimeric rats[J]. Arch Med Sci, 2020, 16(3): 699-706. doi: 10.5114/aoms.2020.94659 [32] 肖艮, 刘静. 茯苓酸调节HMGB1/TLR4信号通路对阿尔兹海默病大鼠神经炎症的影响[J]. 河北医药, 2025, 47(8): 1243-1247. [33] MU P P, LIU Y T, JIANG S M, et al. Glial cell line-derived neurotrophic factor alters lipid composition and protein distribution in MPP+-injured differentiated SH-SY5Y cells[J]. J Cell Physiol, 2020, 235(12): 9347-9360. doi: 10.1002/jcp.29738 [34] LISTA S, GONZÁLEZ-DOMÍNGUEZ R, LÓPEZ-ORTIZ S, et al. Integrative metabolomics science in Alzheimer’s disease: Relevance and future perspectives[J]. Ageing Res Rev, 2023, 89: 101987. doi: 10.1016/j.arr.2023.101987 [35] BHATTI J S, KHULLAR N, MISHRA J, et al. Stem cells in the treatment of Alzheimer’s disease–Promises and pitfalls[J]. Biochim Biophys Acta BBA Mol Basis Dis, 2023, 1869(6): 166712. [36] RAO A, CHEN N, KIM M J, et al. Microglia depletion reduces human neuronal APOE4-related pathologies in a chimeric Alzheimer’s disease model[J]. Cell Stem Cell, 2025, 32(1): 86-104. e7. [37] OYEBODE O D, TULAY P. Mesenchymal stem cells applications in Alzheimer’s disease[J]. Glob Med Genet, 2023, 10(4): 382-387. doi: 10.1055/s-0043-1777087 [38] GIOVANNELLI L, BARI E, JOMMI C, et al. Mesenchymal stem cell secretome and extracellular vesicles for neurodegenerative diseases: Risk-benefit profile and next steps for the market access[J]. Bioact Mater, 2023, 29: 16-35. doi: 10.1016/j.bioactmat.2023.06.013 [39] LIU S, FAN M, XU J X, et al. Exosomes derived from bone-marrow mesenchymal stem cells alleviate cognitive decline in AD-like mice by improving BDNF-related neuropathology[J]. J Neuroinflammation, 2022, 19(1): 35. doi: 10.1186/s12974-022-02393-2 [40] HONG C G, CHEN M L, DUAN R, et al. Transplantation of nasal olfactory mucosa mesenchymal stem cells benefits Alzheimer’s disease[J]. Mol Neurobiol, 2022, 59(12): 7323-7336. doi: 10.1007/s12035-022-03044-6 [41] YU X J, LI Y C, MU X H. Effect of quercetin on PC12 Alzheimer’s disease cell model induced by aβ(25-35)and its mechanism based on Sirtuin1/Nrf2/HO-1 pathway[J]. Biomed Res Int, 2020, 2020: 8210578. doi: 10.1155/2020/8210578 [42] BELL M, ZEMPEL H. SH-SY5Y-derived neurons: a human neuronal model system for investigating TAU sorting and neuronal subtype-specific TAU vulnerability[J]. Rev Neurosci, 2021, 33(1): 1-15. doi: 10.20944/preprints202006.0203.v2 [43] ROYTRAKUL S, JARESITTHIKUNCHAI J, PHAONAKROP N, et al. Secretomic changes of amyloid beta peptides on Alzheimer’s disease related proteins in differentiated human SH-SY5Y neuroblastoma cells[J]. PeerJ, 2024, 12: e17732. doi: 10.7717/peerj.17732 [44] YANAKIEV M, SOPER O, BERG D A, et al. Modelling Alzheimer’s disease using human brain organoids: current progress and challenges[J]. Expert Rev Mol Med, 2022, 25: e3. doi: 10.1017/erm.2022.40 [45] CHEN X W, SUN G Q, TIAN E, et al. Modeling sporadic Alzheimer’s disease in human brain organoids under serum exposure[J]. Adv Sci, 2021, 8(18): 2101462. doi: 10.1002/advs.202101462 [46] ZHANG Y B, TANG C, HE Y, et al. Semaglutide ameliorates Alzheimer’s disease and restores oxytocin in APP/PS1 mice and human brain organoid models[J]. Biomed Pharmacother, 2024, 180: 117540. doi: 10.1016/j.biopha.2024.117540 [47] BHARDWAJ S, KESARI K K, RACHAMALLA M, et al. CRISPR/Cas9 gene editing: New hope for Alzheimer’s disease therapeutics[J]. J Adv Res, 2022, 40: 207-221. doi: 10.1016/j.jare.2021.07.001 [48] FERNANDES T, MELO T, CONDE T, et al. Mapping the lipidome in mitochondria-associated membranes(MAMs)in an in vitro model of Alzheimer’s disease[J]. J Neurochem, 2024, 168(7): 1237-1253. [49] XIONG X M, LI S J, HAN T L, et al. Study of mitophagy and ATP-related metabolomics based on β-amyloid levels in Alzheimer’s disease[J]. Exp Cell Res, 2020, 396(1): 112266. doi: 10.1016/j.yexcr.2020.112266 [50] TRAXLER L, HERDY J R, STEFANONI D, et al. Warburg-like metabolic transformation underlies neuronal degeneration in sporadic Alzheimer’s disease[J]. Cell Metab, 2022, 34(9): 1248-1263. e6. [51] 王辉, 蔡颖, 刘敏, 等. 基于UHPLC-QTOF/MS的细胞代谢组学对丹参和知母防治阿尔茨海默病的药效比较研究[J]. 药学学报, 2021, 56(9): 2394-2402. doi: 10.16438/j.0513-4870.2021-0646 [52] JUNG E S, CHOI H, MOOK-JUNG I. Decoding microglial immunometabolism: a new frontier in Alzheimer’s disease research[J]. Mol Neurodegener, 2025, 20(1): 37. doi: 10.1186/s13024-025-00825-0 [53] QIU Y G, HOU Y, WETZEL L, et al. Cell type-specific master metabolic regulators of Alzheimer’s disease[J]. bioRxiv, 2025. DOI: 10.1101/2025.07.11.664443. [54] ISAAC A, STAN L, HUSSEIN N, Y. Uncovering mechanisms of brain inflammation in Alzheimer’s disease with APOE4: application of single cell-type lipidomics[J]. Ann N Y Acad Sci, 2022, 1518(1): 84-105. doi: 10.1111/nyas.14907 [55] ASADIAN M, CROSLOW S W, TRINKLEIN T J, et al. High-throughput fluorescence-guided sequential single-cell MALDI-ICC mass spectrometry[J]. Anal Chem, 2025, 97(29): 15864-15872. doi: 10.1021/acs.analchem.5c02092 [56] CHEN Y W, HU D J, ZHAO L S, et al. Unraveling metabolic alterations in transgenic mouse model of Alzheimer’s disease using MALDI MS imaging with 4-aminocinnoline-3-carboxamide matrix[J]. Anal Chim Acta, 2022, 1192: 339337. doi: 10.1016/j.aca.2021.339337 [57] CAIRNS J L, HUBER J, LEWEN A, et al. Mass-guided single-cell MALDI imaging of low-mass metabolites reveals cellular activation markers[J]. Adv Sci, 2025, 12(5): 2410506. doi: 10.1002/advs.202410506 -

 点击查看大图
点击查看大图
图(1)
计量
- 文章访问数: 1192
- HTML全文浏览量: 535
- PDF下载量: 4
- 被引次数: 0
