

 下载:
下载:

-
疫苗是当今医学和公共卫生领域最重要的成就之一, 使得全球多种传染病的发病率和死亡率显著降低[1]。自20世纪60年代以来,口服脊髓灰质炎疫苗作为首款口服疫苗,标志着口服疫苗在疫苗开发领域的重要突破[2]。相较于传统注射疫苗,口服疫苗具有无创、无痛、操作简便、患者依从性高、可同时诱导黏膜免疫与全身免疫等显著优势,尤其适用于大规模人群接种[3],因此在疫苗研发领域备受瞩目。抗原经口服后,穿过肠上皮细胞层,被肠道黏膜相关淋巴组织中的抗原递呈细胞识别,引发免疫应答[4]。微褶皱细胞(M 细胞)是一类特化的上皮细胞,承担着“抗原哨兵”的角色, 在黏膜免疫应答中起着关键作用。M细胞数目稀少,但具备卓越的跨上皮转运能力,可高效摄取而不降解腔内抗原物质,将抗原完整地传递至抗原递呈细胞,启动免疫反应。因此,通过靶向M细胞突破黏膜屏障并实现高效递送,可显著提高口服疫苗的免疫效力。
本文围绕M细胞的靶向递送,综述了其生物学特性与递送挑战,归纳了当前主流的靶向策略,并系统介绍了脂质载体、聚合物载体、无机纳米载体与工程化微生物等递送载体的最新研究进展,旨在为新一代口服疫苗及黏膜递送系统的设计开发提供参考。
-
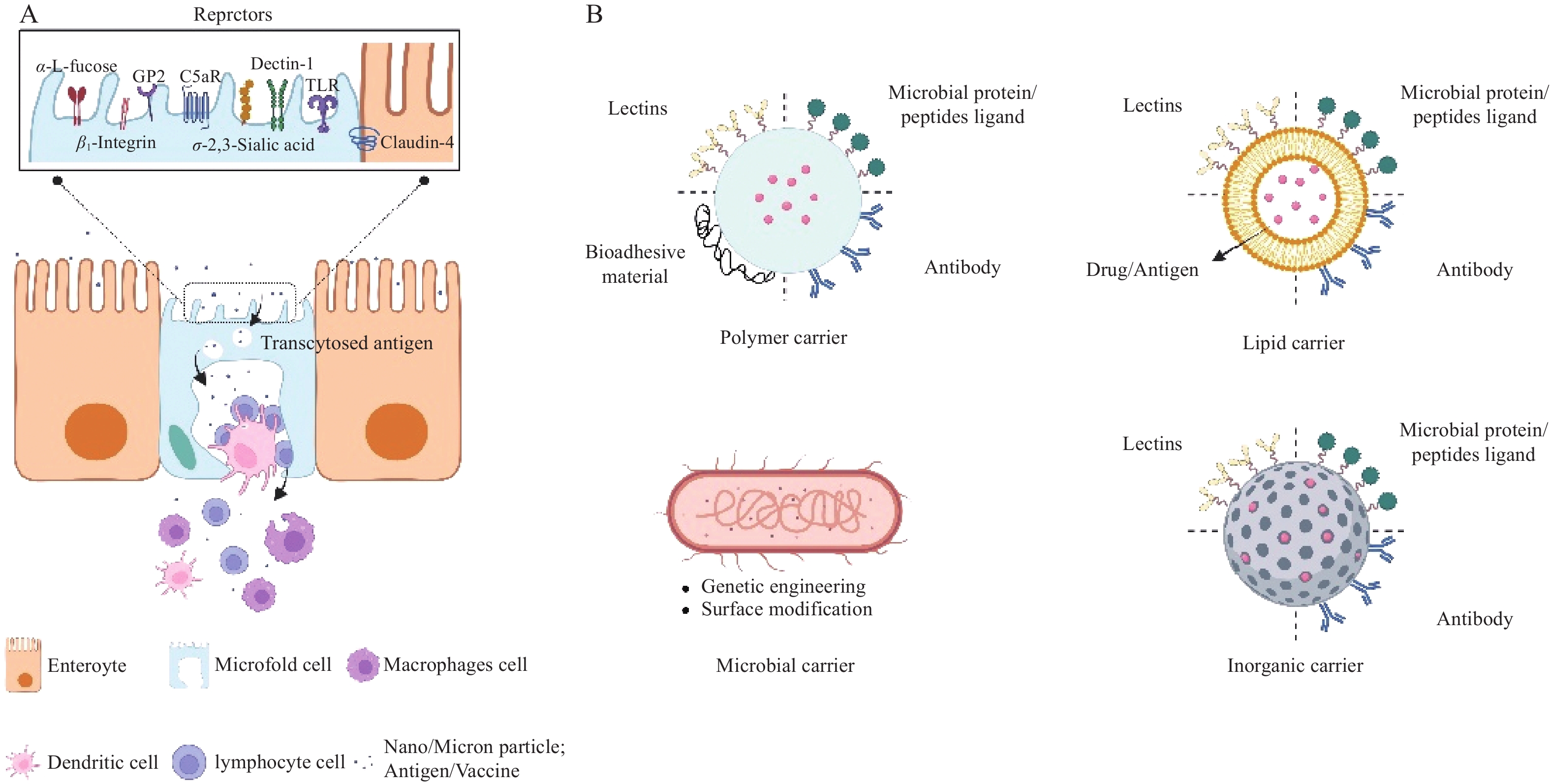
M细胞是一种高度特化的免疫监视上皮细胞,主要位于小肠黏膜派尔集合淋巴结的滤泡相关上皮中。其独特的细胞结构是高效执行抗原摄取与转运的基础,如图1所示。在细胞顶端,M细胞缺乏致密刷状缘和糖萼,微绒毛稀疏且黏液分泌少[5],从而极大增加了与肠腔抗原的接触机会。同时,其顶端表达多种受体也便于主动捕获抗原 [3]。在基底侧,M细胞细胞膜深陷形成 “口袋状”结构,为免疫细胞提供了富集和驻留的场所[5],同时也缩短了抗原胞内转运路径。在胞内,其溶酶体活性低下[6],可有效避免抗原降解,从而将完整抗原快速传递给抗原递呈细胞,高效启动黏膜免疫应答。
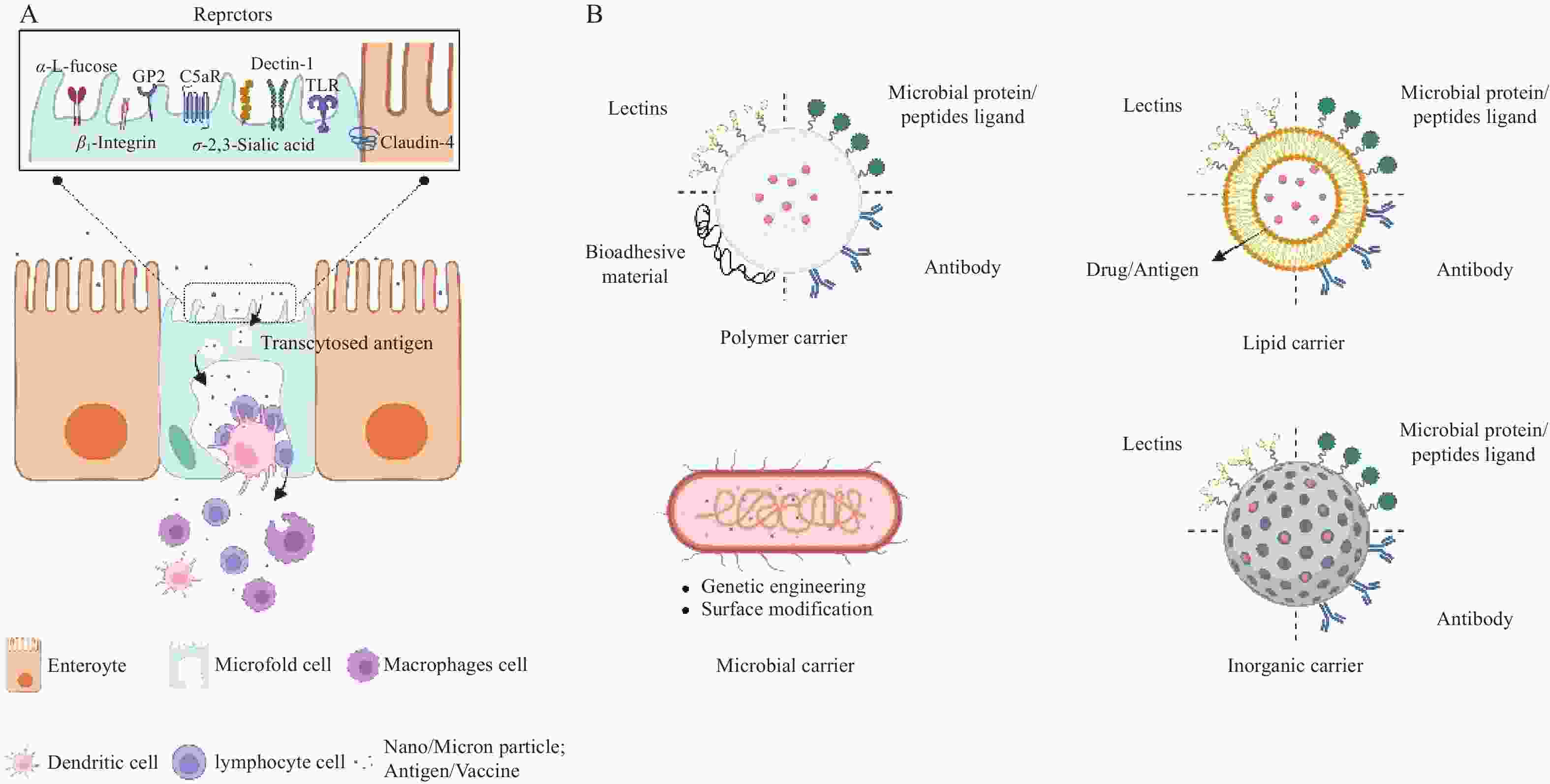
图 1 XXXXXX
M细胞数量稀少,约占1%的肠细胞,但具备卓越的跨上皮转运能力[7]。其抗原摄取的方式分为两种:被动摄取和受体介导主动摄取。被动摄取主要依赖颗粒的理化性质,如粒径较小、呈中性或者负电性的粒子更容易被M细胞吸收[8]。相比之下,主动摄取机制靶向性更强、效率更高,其关键在于利用M细胞表面的特异性或高表达的受体[7],如糖蛋白2(GP2)、补体C5a受体(C5aR )、岩藻糖残基、整合素β1(β1 integrin)、C型凝集素样受体(Dectin-1)、表皮生长因子样重复A结构域(EGF-A)等,如图1所示。这些受体赋予M细胞强大的病原体识别能力,但同时也为病原体的入侵提供了“门户”。基于此,利用仿生技术模拟病原体与M细胞的高亲和力相互作用,从而实现疫苗或活性大分子的靶向递送。
-
尽管M细胞靶向递送潜力巨大,但其开发仍面临着严峻挑战。胃肠道的恶劣环境和消化酶作用会降低疫苗的稳定性和免疫原性[10]。不同肠道区段pH值的变化也可能会进一步干扰疫苗的稳定性。肠道广大的肠表面积、覆盖黏液层以及紧密相连的上皮结构,降低上皮组织的通透性,进一步限制了抗原的有效吸收[9]。再者为了维持对食物和共生菌群的稳态,肠道免疫系统对抗原更可能倾向于免疫耐受而非免疫应答[9],这会削弱疫苗诱导免疫反应的能力,造成免疫原性不足。
同时,M细胞的生物学特性也为其递送研究构成了内在限制。在体内层面,M细胞在肠道上皮中分布稀疏,数目极少,这严重限制了抗原的摄取效率。在体外研究层面,原代M细胞分离与培养困难,目前广泛使用的 Caco-2与Raji细胞共培养模型虽能诱导 “M样细胞”,但仍无法完全模拟人体肠道的复杂生理功能[3]。此外,物种差异问题也不容忽视,许多在小鼠体内有效的靶向配体,如荆豆凝集素(UEA-1),在人体内的特异性亲和力可能不足甚至缺乏相应的结合位点,这也给开发相应的递送技术带来了极大的挑战[3]。
-
M细胞在黏膜免疫应答中起着关键作用,但其稀缺性显著限制了抗原的摄取与递送效率。为克服该瓶颈,利用M细胞表面受体与其特异性配体之间的高亲和力识别机制,通过主动靶向策略增强外源性抗原颗粒或纳米载体的特异性摄取与胞吞转运[11]。目前常应用的M细胞靶向配体与受体主要有以下几类。
-
凝集素是目前研究最为广泛的M细胞靶向配体之一,可通过特异性识别M细胞表面表达的糖基残基实现高效靶向。其中,UEA-1 是一种常用的凝集素,对M细胞表面α-L-岩藻糖残基具有高亲和力,被广泛应用于口服递送系统的靶向修饰[12]。但UEA-1作为一种外源蛋白质,其本身具有潜在的免疫原性和细胞毒性,且在胃肠道中可能被降解,使其应用前景受限。同时人体内M细胞也缺乏对应的受体表达,限制了该凝集素在人体方面的应用[3]。除了UEA-1之外,多种替代性凝集素及其模拟物也被研究用于M细胞的靶向递送[13]。橙黄网孢盘菌凝集素同样可特异性识别α-L-岩藻糖残基,具有与UEA-1相当的M细胞靶向能力,但毒性较低、安全性更优。但其作用机制仍依赖于岩藻糖识别,同样面临相似的种属差异问题。还有凝集素Galectin-9也可实现M细胞的特异性靶向。总体而言,凝集素类配体在M细胞靶向递送中展现出良好前景,但其种属差异性、稳定性及安全性等问题仍需进一步优化。
-
M细胞表面高表达多种模式识别受体及其他特异性膜蛋白[7],成为多种肠道病原体识别并入侵宿主的重要门户。利用病原体与M细胞相互作用的分子机制,模拟其自然侵袭过程,已成为开发高效口服递送系统的一种极具前景的靶向策略。其中,GP2被公认为成熟M细胞的标志性受体,在人和小鼠M细胞中均特异性表达,并可特异性识别革兰氏阴性菌Ⅰ型菌毛上的黏附蛋白FimH,介导如鼠伤寒沙门菌等病原体的捕获与跨膜转运[14]。紧密连接蛋白claudin 4不仅是构成肠上皮细胞间紧密连接的重要蛋白分子,也在M细胞膜表面表达。该蛋白可特异性结合产气荚膜梭菌肠毒素C端30个氨基酸[15]。C5aR同样在M细胞顶侧特异性表达,使其能够响应局部补体激活信号。有研究发现,某些细菌外膜蛋白H是C5aR的靶向配体[16]。此外,M细胞表面表达的Toll样受体4(TLR-4)和TLR-2是革兰氏阴性菌表达的脂多糖和革兰氏阳性菌脂磷壁酸的受体[17]。耶尔森菌的侵袭蛋白通过高亲和力结合M细胞表面的β1 integrin受体,促进细菌内化[18]。其他潜在靶点还包括可被酵母细胞壁β-葡聚糖(β Glus)识别的Dectin-1受体,以及可被病毒σ1蛋白特异性结合的α-2,3-唾液酸受体等[19],进一步拓展了基于受体与配体识别的M细胞靶向策略。
-
除如上提到的两类靶向配体,基于肽序列与特异性抗体的M细胞靶向策略亦取得显著进展。研究表明,精氨酸-甘氨酸-天冬氨酸(RGD)序列可通过识别M细胞表面的β1整合素受体实现有效靶向[20]。利用噬菌体技术筛选获得的多种具有M细胞靶向能力的多肽,如Co-1肽可特异性结合M细胞表面C5aR受体[21]。归巢多肽CKSTHPLSC则通过靶向EGF-A实现M细胞靶向[22]。近期一项报道表明, Li等[23] 通过噬菌体优化筛选获得一种新型D型肽DGPBP-2(1-8),可特异性识别M细胞表面GP2受体,显著增强口服疫苗的黏膜免疫应答。同时靶向M细胞表面标志物的单克隆抗体亦展现出广阔应用前景。例如,针对α(1,2)-岩藻糖受体的mAb NKM16-2-4和GP2受体的anti-GP-2均可有效实现抗原递送至M细胞,并诱导强烈的黏膜免疫应答[24]。研究发现分泌型免疫球蛋白A(sIgA)可通过与Dectin-1受体结合实现M细胞靶向,口服免疫缺陷病毒抗原P24与sIgA复合物后可显著激活肠道黏膜免疫反应[25],另有研究证实分泌型免疫球蛋白M也具备M细胞靶向功能[26]。多肽与抗体类配体的发展为构建高效口服疫苗及M细胞递送系统提供了重要的理论基础。
-
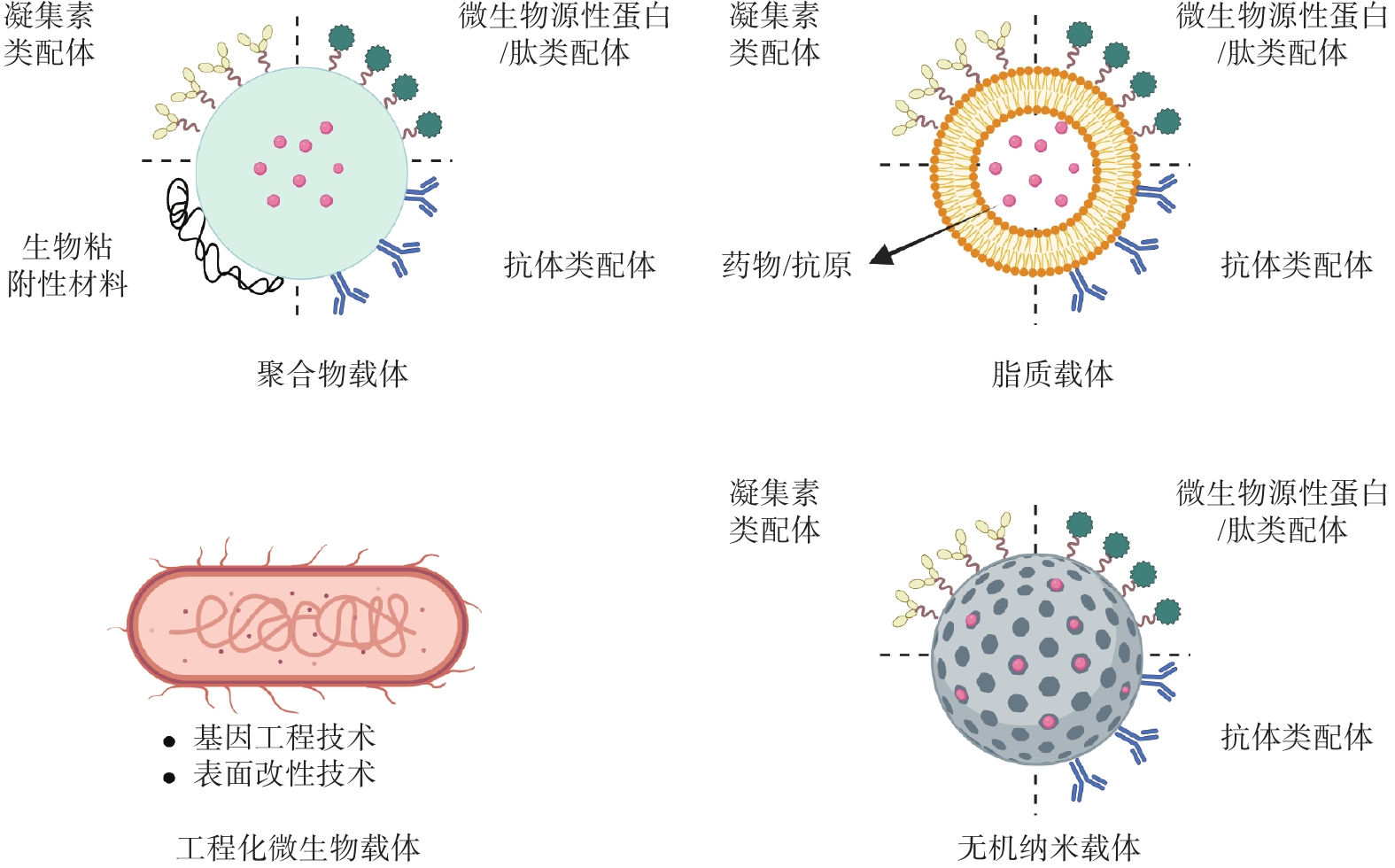
综上,M细胞主动靶向策略的核心在于利用特异性配体实现精准识别。而这些配体必须与递送载体相结合,才能构成完整的靶向递送系统。载体不仅为配体提供了附着位点,更重要的是能够保护负载物、克服生理屏障并控制释放。因此,本章节重点介绍各类递送载体(图2),并展示它们如何通过功能化修饰,将M细胞的靶向从理论转化为实际应用。
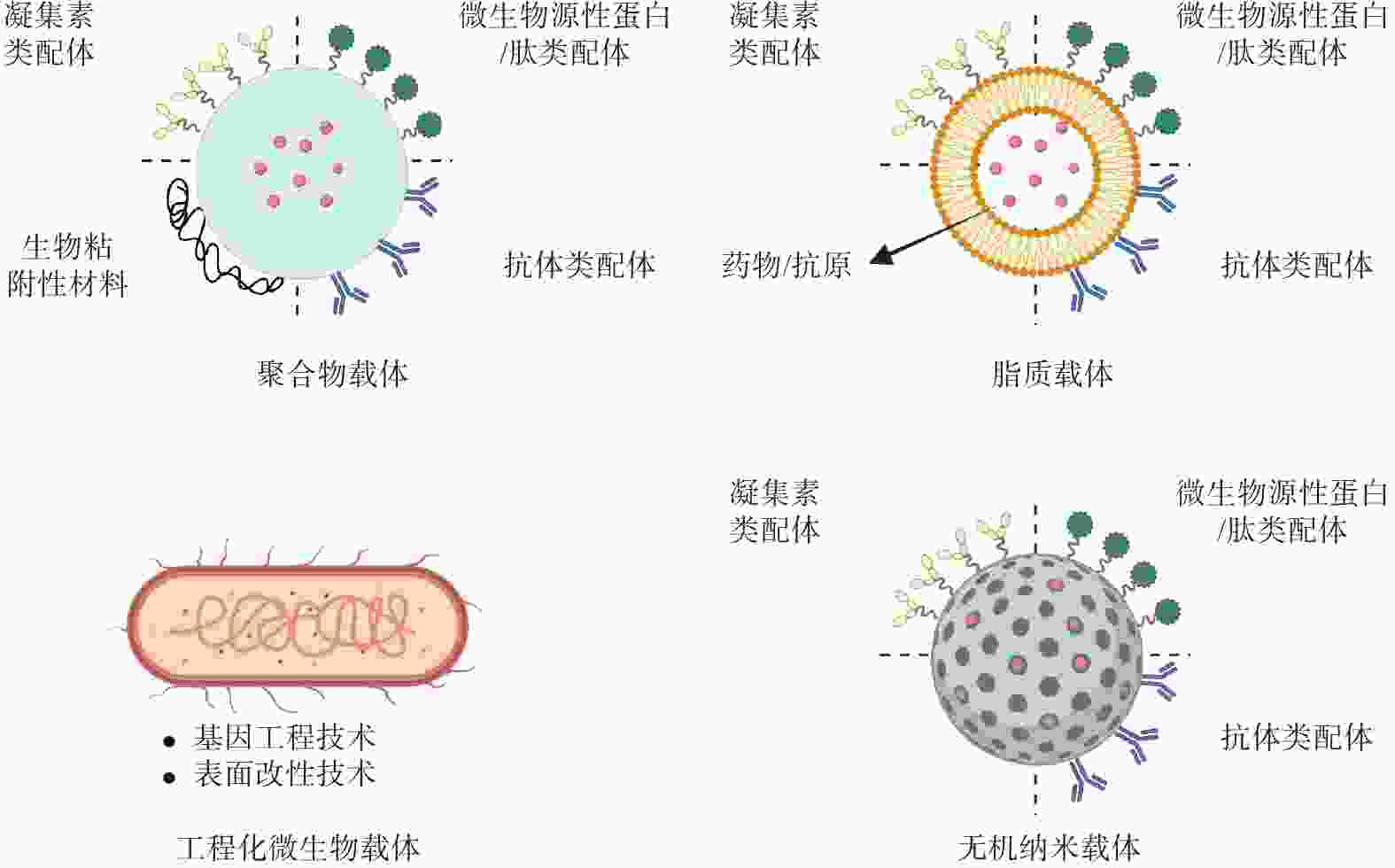
图 2 XXXXXX
-
脂质载体是一类基于天然或人工合成磷脂构建的递送系统,具有制备工艺简便、生物相容性优异、表面功能化修饰灵活以及载药效率高等显著优势。近年来在口服小分子药物、多肽甚至基因的靶向递送领域展现出广阔的应用前景。Luo等[27]报道了一种负载干扰素-γ基因(IFN-γ mRNA)和β Glus修饰的脂质纳米粒口服肿瘤疫苗(IFN-γ mRNA@β Glus-LNPs),口服后通过β Glus特异性识别Dectin-1受体被M细胞摄取,跨越肠道屏障后被内源性巨噬细胞内化运输至肿瘤部位,促进肿瘤部位的巨噬细胞重编程为M1型表型,发挥抗肿瘤作用。此外, Xin等[28]开发了一种脂质-聚合物杂化纳米粒口服疫苗递送平台(mPOD),其表面修饰了一种来源于革兰氏阴性和革兰氏阳性细菌肽聚糖分解产物的二肽,通过模拟细菌表面结构靶向M细胞表面的肠道二肽转运体。该策略有效促进了抗原递送至黏膜免疫系统,成功激活了抗白血病免疫反应。近期研究[29]还报道了一种新型口服紫杉醇递送系统,该系统可在肠道原位生包裹紫杉醇纳米乳液油滴(DA-β Glus/ODs/PTX),口服后通过β Glus靶向M细胞,并借助内源性巨噬细胞将紫杉醇高效递送至肿瘤微环境,触发肿瘤细胞免疫原性死亡,实现协同增强化疗疗效与肿瘤免疫疗法。
-
除了脂质载体,由天然或者合成的大分子材料所组成的聚合物载体在M细胞靶向递送活性成分方面同样具有广泛的应用。例如,Wang等[30]构建了一种口服肿瘤疫苗(CpG@OVAHBc@Dex),是由水凝胶负载一种表面嵌合肿瘤抗原并内部装载佐剂核苷酸的乙肝病毒核心抗原的病毒样颗粒组成的。口服后有效保护疫苗成分免受胃酸和酶的降解,并在肠道中定向释放。病毒样颗粒通过模仿病毒结构增强黏膜表面的黏附性,促进了M细胞的摄取和抗原递呈细胞的递呈,从而激活了黏膜和全身免疫反应。而另外一项研究中,Li等[23]设计了一种由GP2靶向肽偶联酵母胶囊组成的口服疫苗平台(GP2-YCs),通过靶向肽穿过M细胞和β Glus增加树突状细胞内吞以实现级联靶向,可有效地激活全身和黏膜免疫反应。在其他方面,Sung团队[31]设计了多种β Glus功能化的聚合物纳米粒,其中一种是口服前药递送系统(β Glus-CTX NPs),该系统是由β Glus与抗生素头孢噻肟通过活性氧响应键连接。口服后利用β Glus跨越肠道屏障,由内源性巨噬细胞携带通过淋巴和循环系统,穿过血脑屏障到达炎症脑组织,在高浓度活性氧作用下释放活性药物,以实现细菌性脑膜炎的口服精准治疗。
-
无机递送系统因其结构和功能多样性,如形状大小可调、光学和磁性特性、比表面积高和表面易于改性等,被广泛用于生物医药等领域,特别是在药物靶向递送、影像诊断和药物协同治疗方面具有广阔的前景。在一项研究[32]报道了一种口服基于天然免疫介导的巨噬细胞靶向纳米平台(β Glus-R848/Ag2Te NPs)。该纳米平台利用β Glus穿过M细胞并靶向巨噬细胞,借助其实现肿瘤递送,并在肿瘤微环境的活性氧信号触发下,释放免疫调节剂R848,将巨噬细胞重编程为抗肿瘤作用的M1型表型。同时,整合的Ag2Te量子点提供深层组织近红外Ⅱ区成像,实现实时监测肿瘤进展和免疫治疗响应。
-
随着生物技术和合成生物学的快速发展,利用微生物作为活体或生物衍生载体在药物、疫苗和治疗性蛋白的递送方面同样具有广泛的应用。近期研究[33]报道了一种以双歧杆菌为载体的口服疫苗(Bif-OVA-Ocur)。该载体是通过层层沉积的方式将肿瘤抗原肽OVA257-264与一种氧化多糖包覆在双歧杆菌表面。口服后,Bif-OVA-Ocur可有效地定向到肠道中的派尔集合淋巴结,通过多糖靶向M细胞的Dectin-1受体,并进一步转运给抗原递呈细胞,随即诱导强效的全身和黏膜免疫反应。在另一项报道中,Chen等[34]开发了一种工程化酵母疫苗(YVm),通过基因工程的手段将SARS-CoV-2受体结合域锚定在酵母表面,并结合酵母自身的天然佐剂和M细胞靶向功能,口服后可显著激活免疫反应。此外,Li等[35]通过设计了一种基于基因编辑与超声响应的工程化大肠杆菌的口服纳米疫苗——E.coli(AH1-CDA-Co1)@iPDA,口服后,利用聚多巴胺涂层保护工程菌并延长肠道驻留。在超声触发下,工程菌分泌携带肿瘤抗原AH1、STING激动剂CDA及M细胞靶向肽Co1的细菌外膜囊泡,作为结肠癌原位疫苗发挥作用。工程菌的外膜囊泡通过靶向M细胞,穿透肠道上皮屏障,激活TLR-4和环鸟嘌呤-腺嘌呤核苷酸合成酶-干扰素基因刺激蛋白天然免疫信号通路,引发肿瘤抗原特异性免疫反应,显著抑制肿瘤生长并延长生存期。该研究还展示了基因工程与超声调控技术在精准治疗中的巨大潜力。
-
口服疫苗兼具无创、便捷、易于大规模分发的特点,并能诱导黏膜和全身双重免疫,优势独特。然而,口服递送需克服胃肠道严苛环境、黏液和上皮屏障的物理阻隔,以及易诱导免疫耐受等多重障碍。M细胞作为黏膜免疫的“门户”, 是实现主动靶向的理想靶点。虽然其数量稀少,但可利用其表面特异性或高表达的多类受体,通过“配体-受体”选择性识别机制进行精准递送。近二十年来,随着对M细胞作用机制理解的深入和靶向配体的不断发现,相关递送技术取得了显著进展,开发出了多种新型递送系统。其中,β-葡聚糖因兼具天然M细胞靶向与免疫调节双重功能而备受关注,基于此开发的口服递送系统已广泛应用于疫苗递送、肿瘤免疫治疗等领域。尽管M细胞靶向递送技术尚未实现临床应用,但其凭借对黏膜屏障的精准“破壁”能力,有望为口服疫苗及生物大分子药物的研发带来革命性的突破。
Progress on M cells-targeted oral vaccine delivery systems
-
摘要: 口服疫苗因兼具无创、便捷、可诱导黏膜与全身双重免疫等优势,成为疫苗研发的重要方向。微褶皱细胞(M细胞),是一类特化的免疫监视上皮细胞,数量虽稀少,但具备高效摄取和转运抗原的能力,是突破口服疫苗递送瓶颈的关键靶点。本文综述肠道M细胞的形态结构、摄取机制与表面受体特征,分析当前疫苗经口服靶向M细胞递送所面临的多项挑战,总结基于受体与配体特异性识别机制的靶向策略进展,同时归纳递送载体当前在M细胞靶向口服递送方面的设计与应用。以期为将来针对肠道M细胞的黏膜靶向递送系统的研究和应用提供借鉴。Abstract: Oral vaccines, owing to their non-invasive nature, ease of administration, and ability to elicit both mucosal and systemic immune responses, represent a pivotal direction in vaccine development. Microfold cells (M cells), a specialized subset of immune-surveillance epithelial cells, though sparsely distributed, possess exceptional capacity for antigen uptake and transcytosis, rendering them a critical target for overcoming the bottlenecks of oral vaccine delivery. The morphological structure, antigen uptake mechanisms, and surface receptor characteristics of intestinal M cells were summarized in this paper. The multifaceted challenges currently impeding M cell-targeted oral delivery and recent advances in targeting strategies based on receptor–ligand specific recognition were analyzed. The design and application of delivery carriers tailored for M cell targeting were reviewed. This study aimed to provide insights and guidance for future research and development of M cell-targeted mucosal delivery systems in the intestine.
-
Key words:
- oral vaccine /
- microfold cell /
- targeted delivery /
- mucosal immunity
-
[1] SHATTOCK A J, JOHNSON H C, SIM S Y, et al. Contribution of vaccination to improved survival and health: modelling 50 years of the Expanded Programme on Immunization[J]. Lancet, 2024, 403(10441): 2307-2316. doi: 10.1016/S0140-6736(24)00850-X [2] PARKER E P, MOLODECKY N A, PONS-SALORT M, et al. Impact of inactivated poliovirus vaccine on mucosal immunity: implications for the polio eradication endgame[J]. Expert Rev Vaccines, 2015, 14(8): 1113-1123. doi: 10.1586/14760584.2015.1052800 [3] ISLAM M A, FIRDOUS J, BADRUDDOZA A Z M, et al. M cell targeting engineered biomaterials for effective vaccination[J]. Biomaterials, 2019, 192: 75-94. doi: 10.1016/j.biomaterials.2018.10.041 [4] DU G S, QIN M, SUN X. Recent progress in application of nanovaccines for enhancing mucosal immune responses[J]. Acta Pharm Sin B, 2023, 13(6): 2334-2345. doi: 10.1016/j.apsb.2022.08.010 [5] KUNISAWA J, KURASHIMA Y, KIYONO H. Gut-associated lymphoid tissues for the development of oral vaccines[J]. Adv Drug Deliv Rev, 2012, 64(6): 523-530. doi: 10.1016/j.addr.2011.07.003 [6] LELOUARD H, FALLET M, DE BOVIS B, et al. Peyer’s patch dendritic cells sample antigens by extending dendrites through M cell-specific transcellular pores[J]. Gastroenterology, 2012, 142(3): 592-601. e3. [7] XU Y N, SHRESTHA N, PRÉAT V, et al. Overcoming the intestinal barrier: a look into targeting approaches for improved oral drug delivery systems[J]. J Control Release, 2020, 322: 486-508. doi: 10.1016/j.jconrel.2020.04.006 [8] DES RIEUX A, RAGNARSSON E G E, GULLBERG E, et al. Transport of nanoparticles across an in vitro model of the human intestinal follicle associated epithelium[J]. Eur J Pharm Sci, 2005, 25(4-5): 455-465. doi: 10.1016/j.ejps.2005.04.015 [9] ZHANG Y D, LI M, DU G S, et al. Advancedoral vaccine delivery strategies for improving the immunity[J]. Adv Drug Deliv Rev, 2021, 177: 113928. doi: 10.1016/j.addr.2021.113928 [10] MIAO Y B, LIN Y J, CHEN K H, et al. Engineering nano- and microparticles as oral delivery vehicles to promote intestinal lymphatic drug transport[J]. Adv Mater, 2021, 33(51): e2104139. doi: 10.1002/adma.202104139 [11] 16] LONGET S, LUNDAHL M L E, LAVELLE E C. Targeted strategies for mucosal vaccination[J]. Bioconjug Chem, 2018, 29(3): 613-623. doi: 10.1021/acs.bioconjchem.7b00738 [12] CLARK M A, BLAIR H, LIANG L, et al. Targeting polymerised liposome vaccine carriers to intestinal M cells[J]. Vaccine, 2001, 20(1-2): 208-217. doi: 10.1016/S0264-410X(01)00258-4 [13] YEBOAH K G, AKANDE J, ADDO R T, et al. In vitro and ex vivo characterization of lectin-labeled Mycobacterium tuberculosis antigen-containing microspheres for enhanced oral delivery[J]. J Drug Target, 2014, 22(1): 34-47. doi: 10.3109/1061186X.2013.833206 [14] HASE K, KAWANO K, NOCHI T, et al. Uptake through glycoprotein 2 of FimH(+)bacteria by M cells initiates mucosal immune response[J]. Nature, 2009, 462(7270): 226-230. doi: 10.1038/nature08529 [15] LING J, LIAO H L, CLARK R, et al. Structural constraints for the binding of short peptides to claudin-4 revealed by surface plasmon resonance[J]. J Biol Chem, 2008, 283(45): 30585-30595. doi: 10.1074/jbc.M803548200 [16] KIM S H, YANG I Y, JANG S H, et al. C5a receptor-targeting ligand-mediated delivery of dengue virus antigen to M cells evokes antigen-specific systemic and mucosal immune responses in oral immunization[J]. Microbes Infect, 2013, 15(13): 895-902. doi: 10.1016/j.micinf.2013.07.006 [17] AKASHI S, SAITOH S I, WAKABAYASHI Y, et al. Lipopolysaccharide interaction with cell surface Toll-like receptor 4-MD-2: higher affinity than that with MD-2 or CD14[J]. J Exp Med, 2003, 198(7): 1035-1042. [18] CLARK M A, HIRST B H, JEPSON M A. M-cell surface beta1 integrin expression and invasin-mediated targeting of Yersinia pseudotuberculosis to mouse Peyer’s patch M cells[J]. Infect Immun, 1998, 66(3): 1237-1243. doi: 10.1128/IAI.66.3.1237-1243.1998 [19] MIAO Y B, CHEN K H, CHEN C T, et al. A noninvasive gut-to-brain oral drug delivery system for treating brain tumors[J]. Adv Mater, 2021, 33(34): e2100701. doi: 10.1002/adma.202100701 [20] HUMPHRIES J D, BYRON A, HUMPHRIES M J. Integrin ligands at a glance[J]. J Cell Sci, 2006, 119(Pt 19): 3901-3903. [21] KIM S H, JUNG D I, YANG I Y, et al. Application of an M-cell-targeting ligand for oral vaccination induces efficient systemic and mucosal immune responses against a viral antigen[J]. Int Immunol, 2013, 25(11): 623-632. doi: 10.1093/intimm/dxt029 [22] YOO M K, KANG S K, CHOI J H, et al. Targeted delivery of chitosan nanoparticles to Peyer’s patch using M cell-homing peptide selected by phage display technique[J]. Biomaterials, 2010, 31(30): 7738-7747. doi: 10.1016/j.biomaterials.2010.06.059 [23] LI W Q, YANG X, WANG M F, et al. Development of a cascade-targeting oral vaccine via glycoprotein 2 on intestinal microfold cells for cancer immunotherapy[J]. Nano Lett, 2025, 25(13): 5165-5173. doi: 10.1021/acs.nanolett.4c06123 [24] SHIMA H, WATANABE T, FUKUDA S, et al. A novel mucosal vaccine targeting Peyer’s patch M cells induces protective antigen-specific IgA responses[J]. Int Immunol, 2014, 26(11): 619-625. doi: 10.1093/intimm/dxu061 [25] ROCHEREAU N, PAVOT V, VERRIER B, et al. Delivery of antigen to nasal-associated lymphoid tissue microfold cells through secretory IgA targeting local dendritic cells confers protective immunity[J]. J Allergy Clin Immunol, 2016, 137(1): 214-222. e2. [26] ROCHEREAU N, MICHAUD E, WAECKEL L, et al. Essential role of TOSO/FAIM3 in intestinal IgM reverse transcytosis[J]. Cell Rep, 2021, 37(7): 110006. doi: 10.1016/j.celrep.2021.110006 [27] LUO P K, CHANG W N, PENG S Y, et al. Endogenous macrophages as “Trojan horses” for targeted oral delivery of mRNA-encoded cytokines in tumor microenvironment immunotherapy[J]. Biomaterials, 2026, 325: 123620. doi: 10.1016/j.biomaterials.2025.123620 [28] XIN X F, WU D, ZHAO P B, et al. Catch-to-amplify nanoparticles with bacteria surface for sequential mucosal immune activation for acute myeloid leukemia therapy[J]. ACS Nano, 2025, 19(15): 14661-14679. doi: 10.1021/acsnano.4c08515 [29] NGUYEN N, HOANG T M, HUANG T, et al. Macrophage-hitchhiked, effervescence-induced nanoemulsions for enhanced oral chemotherapy and immunotherapy: Impact on absorption route[J]. Biomaterials, 2025, 316: 123019. doi: 10.1016/j.biomaterials.2024.123019 [30] WANG C F, TANG H B, DUAN Y F, et al. Oral biomimetic virus vaccine hydrogel for robust abscopal antitumour efficacy[J]. J Colloid Interface Sci, 2024, 674: 92-107. doi: 10.1016/j.jcis.2024.06.102 [31] NGUYEN V K, NGUYEN N, LI Z C, et al. Inflammation-activated endogenous macrophage-mediated prodrug delivery system overcoming biological barriers for enhanced oral meningitis therapy[J]. Adv Funct Mater, 2024, 34(32): 2401570. doi: 10.1002/adfm.202401570 [32] SIBORO P Y, NGUYEN N, LO S K, et al. Innate immunity-guided macrophage-homing nanoplatform for oral tumor immunotherapy and real-time deep-tissue imaging in pre-clinical models[J]. Adv Mater, 2025, 37(40): e07607. doi: 10.1002/adma.202507607 [33] CHEN Z, QIN Y T, LI Q R, et al. Layer-by-layer deposition of antigen peptides on Bifidobacterium for subintestinal lymphatic system-guided personalized tumor immunotherapy[J]. Adv Mater, 2025, 37(26): e2503571. doi: 10.1002/adma.202503571 [34] CHEN X N, SHI T F, CHEN F M, et al. Orally antigen-engineered yeast vaccine elicits robust intestinal mucosal immunity[J]. ACS Nano, 2025, 19(11): 10841-10853. doi: 10.1021/acsnano.4c14690 [35] LI J X, YANG R Q, YUAN Y H, et al. An oral nanovaccine secreted by genetically engineered and ultrasound-responsive bacteria for colon cancer immunotherapy[J]. Adv Funct Mater, 2025, 35(6): 2414994. doi: 10.1002/adfm.202414994 -

 点击查看大图
点击查看大图
图(2)
计量
- 文章访问数: 249
- HTML全文浏览量: 89
- PDF下载量: 1
- 被引次数: 0
