

 下载:
下载:

-
根据国际癌症研究机构(IARC)发布的2022年全球癌症报告(GLOBOCAN)[1]:在全球范围内,结直肠癌(CRC)发病率位居男性恶性肿瘤的第3位,女性恶性肿瘤的第2位;2022年,全球CRC预计有192.6万新发病例和90.4万死亡病例。中国癌症报告2024显示[2]:2022年,我国CRC发病率位居全国恶性肿瘤第2位(10.74%),病死率位居全国恶性肿瘤第4位(8.42%),且近十几年来我国CRC的发病率和病死率均呈持续上升趋势。
胆汁酸(BAs)是生命活动的重要调节因子,可通过BAs受体参与调节葡萄糖、脂质和能量代谢,并与肠道激素、微生物群和能量平衡密切相关[3]。除此之外,BAs受体还参与免疫调节,BAs受体的免疫调节作用可能与CRC的发生、发展有关。BAs水平在很大程度上是通过BAs受体在转录水平上调节的,BAs受体在BAs的合成、运输和解毒中起关键作用。次级BAs被认为是CRC的促癌因子,其水平的异常升高可通过细胞凋亡、DNA氧化损伤和细胞增殖等机制,促进炎症和CRC的发生。本文拟对BAs受体的免疫调节功能及其在CRC发生、发展中的作用进行综述。
-
BAs的合成包括常规途径和补偿途径,至少有17种酶参与全过程。常规途径是BAs的主要合成方式,在正常生理条件下至少有70%的BAs由该途径产生(图1)。有报道称,在肝脏疾病患者体内,补偿途径成为主要的BAs合成途径[4]。常规途径的第一步是胆固醇经胆固醇7α-羟化酶(CYP7A1)催化生成7α-羟基胆固醇,7α-羟基胆固醇进一步催化生成胆酸(CA)和鹅去氧胆酸(CDCA)。替代途径由线粒体内膜上的胆固醇27-羟化酶(CYP27A1)催化胆固醇27生成27-羟化胆固醇开始,27-羟化胆固醇再经氧化胆固醇7α-羟化酶(CYP7B1)催化,形成7α-羟基化中间体,该中间体再经固醇环修饰、侧链氧化和截短等一系列步骤,最终生成CDCA。值得注意的是,补偿途径主要产生CDCA,而常规途径可以同时产生CA和CDCA,CA与CDCA的比例由合成CA所必需的甾醇-12α-羟化酶(CYP8B1)决定。BAs稳态的改变和消除涉及几种代谢途径,主要包括酰胺化、羟基化、硫酸化和葡萄糖醛酸化途径[5]。下面分别对这几种代谢途径作一介绍。
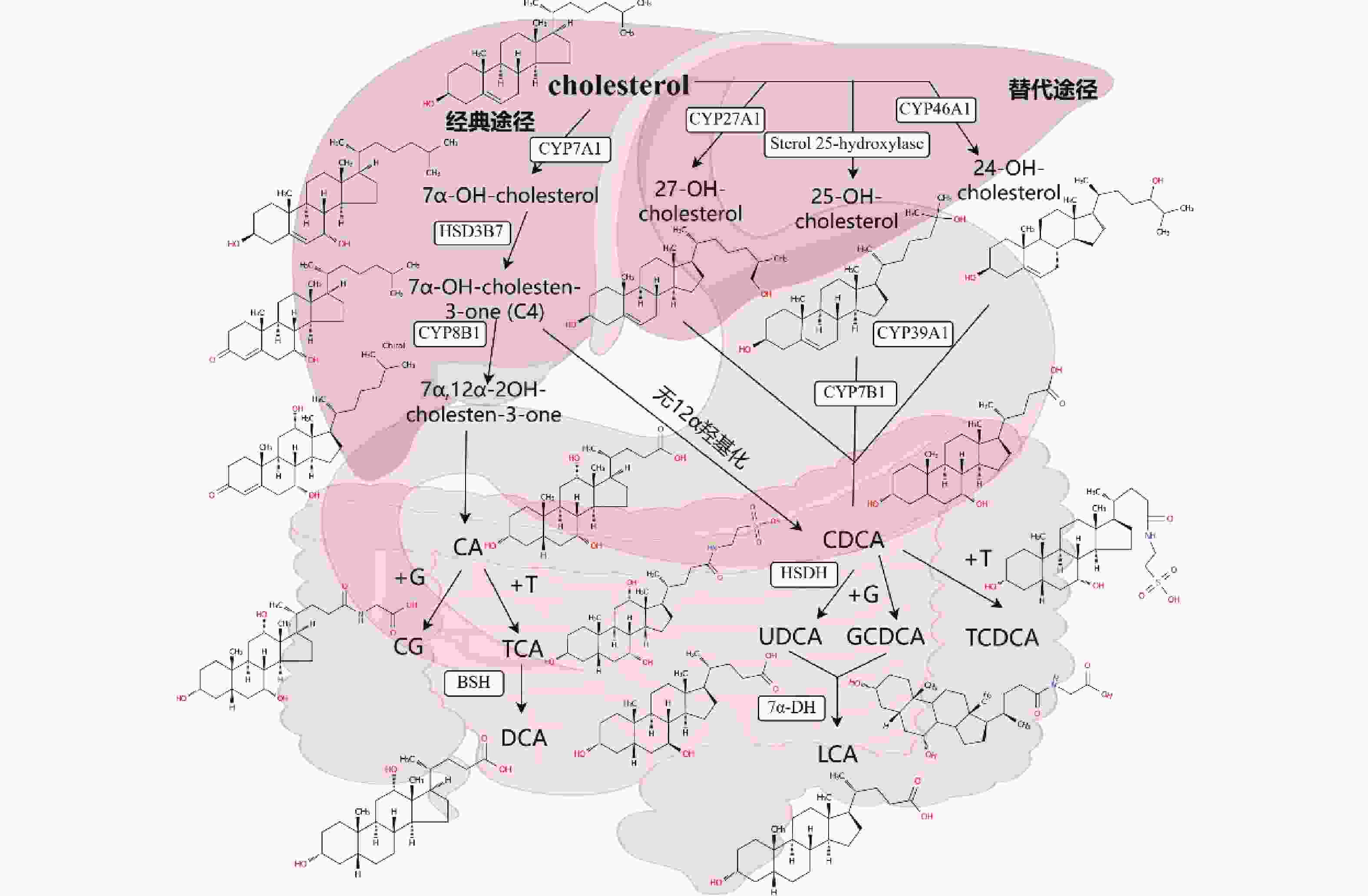
图 1 胆汁酸的合成与代谢
-
在人体中,非结合型BAs主要与甘氨酸发生酰胺化反应[6],研究发现,BAs通过BAs-辅酶A (CoA)/氨基酸N酰基转移酶在过氧化物酶体中与牛磺酸或甘氨酸发生酰胺化反应。Greim等[7]发现,新生儿肝内胆汁淤积症(NICCD)患者血浆中结合型BAs[如甘氨鹅去氧胆酸(GCDCA)]的水平升高,且GCDCA是胆汁淤积症患者血清和胆汁中最丰富的BAs,提示BAs的酰胺化反应可能是机体的一种防御机制。然而,Higuchi等[8]发现,大鼠、人类肝细胞或肝细胞系经高浓度(100 mol/L)GCDCA处理后,会出现严重的线粒体功能障碍和凋亡,表明BAs酰胺化作用的过度上调,可能对机体造成损伤。
BAs的疏水性与甾体环上的羟基数目呈反比。进一步研究发现,BAs的疏水性还取决于羟基的位置和立体化学性质,与C7α-羟基相比,C7β-羟基、C6α-羟基和C6β-羟基更加亲水。一般来说,疏水性BAs[石胆酸(LCA)和去氧胆酸(DCA)]同亲水性BAs[CA、熊去氧胆酸(UDCA)、鼠胆酸(MCA)和猪胆酸(HCA)]相比,具有更强的细胞毒性。Thakare等[6]发现,在人类以及8种动物(包括小型猪、兔子、仓鼠、大鼠、小鼠、黑猩猩、猴子、比格犬)中,三羟基BAs在尿液中的比例(12%~94%)高于血浆(4%~75%),而单羟基BAs在尿液中的比例(0.01%~19%)低于血浆(0.06%~52%),表明羟基化反应有促进BAs经尿液排泄的作用。
-
在人尿液中85%的BAs以硫酸盐形式存在,而在血浆中仅有32%的BAs以硫酸盐形式存在,表明硫酸化反应也可能有促进BAs经尿液排泄的作用。BAs硫酸化程度与其羟基数目成反比,如单羟基BAs LCA几乎是100%硫酸化,而三羟基BAs的硫酸化程度则较低。随着BAs羟基数目的减少,BAs的硫酸化程度升高,说明硫酸化反应对毒性BAs有解毒作用。Huang等[9]认为,BAs通过硫酸化反应,增加了水溶性,降低了其在肠道中的吸收,从而促进BAs经尿液、粪便排泄,并通过增加临界胶束浓度(CMC),降低其细胞毒性。在人体中,BAs葡萄糖醛酸化的程度较低(<12%),以酰基葡萄糖醛酸化反应为主。与硫酸化反应相似,BAs葡萄糖醛酸化程度与羟基数目成反比。Huang等[9]认为,硫酸化和葡萄糖醛酸化是BAs解毒的重要途径。
-
核受体(NR)是配体激活的转录因子,在NR超家族中,至少有5个成员可以作为BAs受体:法尼醇受体(FXR)、孕烷X受体(PXR)、组成型雄甾烷受体(CAR)、维生素D受体(VDR)和肝X受体α(LXRα)。此外,G蛋白偶联受体(TGR5)和鞘氨醇-1-磷酸2型受体(S1PR2)也是BAs的受体[10]。其中,FXR、TGR5、CAR、PXR、VDR具有免疫调节功能,在结直肠癌的发生、发展中发挥重要作用。下面分别对这5种BAs受体的免疫调节功能作一介绍。
-
FXR被认为是结肠炎症和结直肠癌病因学的重要因素。与对照组相比,在FXR基因敲除的小鼠中,实验诱导的肠道炎症严重程度明显增加[11]。在小鼠中,胆管结扎(BDL)和阻断胆汁流向肠道会导致细菌过度生长和黏膜损伤,而在没有FXR的情况下,这些损伤会加剧[12]。研究发现,FXR可通过介导诱导型一氧化氮合酶(iNOS)和IF-18等的表达,抑制细菌的生长[13]。此外,研究表明,CDCA可通过FXR诱导上皮细胞产生抗菌肽,这种机制可能与BAs的直接抑菌作用相结合,防止小肠细菌过度生长[14]。Gadaleta等[15]研究发现,奥贝胆酸(OCA)对葡聚糖硫酸钠(DSS)和2,4,6-三硝基苯磺酸(TNBS)诱导的小鼠结肠炎有保护作用,他们还发现在肠易激综合征患者(IBD)中,经FXR激动剂激活后的FXR,可以抑制单核细胞分泌肿瘤坏死因子α(TNF-α)。这些结果提示FXR激动剂在肠易激综合征等炎症性疾病中有潜在的治疗作用。
早期研究发现[16],TGR5基因敲除的小鼠对TNBS诱导的结肠炎更易感,提示TGR5具有抗炎作用。在经脂多糖(LPS)处理的人类原代巨噬细胞中,发现牛磺石胆酸(TLCA)和其他TGR5配体可促进抗炎细胞因子IL-10的表达,并抑制促炎细胞因子IL-12的表达[17]。Biagioli等[18]研究发现,当TGR5激活后,可引起肠道巨噬细胞M1/M2表型的改变,并降低促炎细胞因子水平,其机制是通过升高环磷腺苷(cAMP)和激活环磷腺苷效应元件结合蛋白(CREB),竞争性干扰促炎调节因子核因子-kB(NF-kB)。牛磺鹅去氧胆酸(TCDCA)和另一种TGR5激动剂[苄基-2-酮-6甲基-4-(2-噻吩)-1,2,3,4-四氢嘧啶-5-羧酸盐]可通过影响人单核细胞向树突状细胞的分化,降低促炎细胞因子IL-12的水平,其效应也是通过cAMP信号介导的[19]。
-
Xie等[20]研究发现,PXR经LCA激活后,可以诱导解毒酶系CYP3A4的表达,进而促进BAs的解毒。肠道PXR可通过抑制核因子-kB和炎症细胞因子的表达,对炎症性肠病有保护作用。另外,PXR和CAR通过调节BAs转运、解毒的相关基因,如BSEP、UGT、MRP3和SULT,对肝脏有保护作用。PXR和CAR的下调均可导致UGT活性、SULT2A1蛋白表达降低,其中,SULT2A1介导的硫酸化反应对BAs的解毒是必要的,可以有效避免结肠癌的发生。然而,有研究发现[21],PXR基因敲除小鼠对LCA的肝毒性具有抗性。因此,需要进一步研究PXR在BAs解毒中的作用。
与PXR类似,VDR经维生素D和LCA激活后,促进解毒酶系CYP3A的转录,从而代谢毒性BAs和其他毒素。肠道VDR虽然可以通过FGF15途径抑制BAs合成,但其主要作用可能是促进BAs解毒。VDR也被证明参与调节免疫反应,如UDCA可通过VDR诱导上皮细胞产生抗菌肽[22]。另外,VDR缺陷小鼠模型的代谢组学研究显示,初级、次级BAs的生成增加与炎症性肠病和结直肠癌的发生有关[23]。研究发现,益生菌对VDR的调节可以抑制肠道炎症和癌变[24]。Wu等[25]研究发现,摄入益生菌鼠李糖乳杆菌GG(LGG)和植物乳杆菌(LP)可以增加肠上皮细胞中VDR的表达。有报道称[26],与对照组相比,结直肠癌组织中VDR基因显著下调。
-
结直肠腺瘤被认为是结直肠癌的一种癌前病变[27]。Yachida等[28]发现,与健康对照组相比,多发性息肉状腺瘤(不包括锯齿状病变)和(或)黏膜内癌患者粪便中DCA的水平明显增加。Brown等[29]发现,与邻近黏膜相比,结直肠癌患者肿瘤组织中DCA的水平降低。同样,多项研究发现,与健康对照组相比,CRC患者粪便中DCA的水平较低[30-32]。但有报道称[33],DCA水平在结直肠癌高发病率人群粪便中增加。对于DCA而言,不同研究结果之间存在争议,进一步扩大样本量可能阐明DCA在结直肠癌发生和发展中的作用。
Williams等[34]发现,结直肠癌患者肿瘤组织中7-酮-DCA、12-酮-DCA和7-羟基-3-氧-胆烷酸的水平上调。进一步分析发现,上述BAs水平的升高只在转移性结直肠癌患者中观察到,原发性结直肠癌组织中这些BAs的水平几乎没有变化,因此这些BAs可能是结直肠癌转移进展和晚期的生物标志物。另外,Chetwynd等[35]发现,在结直肠癌患者粪便中,2种酮结构BAs浓度显著升高,但未被识别,低丰度酮BAs在结直肠癌中作用有待进一步研究。近年来代谢组学研究发现,与对照组相比,结直肠癌以及息肉或腺瘤患者体内BAs水平存在差异(表1),这些差异BAs可能作为结直肠癌诊断的潜在生物标志物。
表 1 结直肠癌、息肉或腺瘤患者体内胆汁酸的变化情况
病例/对照 样本 胆汁酸变化情况 文献 CA GCA TCA CDCA GCDCA LCA DCA 7-酮DCA 12-酮DCA 7-OH-3O胆烷酸 CRC组织(n=10);
良性组织(n=9)组织 ↑a ↑a ↑a [34] 结肠腺瘤(n=59);
CRC(n=56,Ⅰ~Ⅳ期各14);
健康对照(n=55)血清 ↓b [36] CRC(n=28);
息肉(n=44);
健康对照(n=55)血清 ↓c ↑d [37] CRC组织(n=17);
邻近黏膜(n=17)组织 ↓a ↓a [32] 结肠腺瘤(n=10);
健康对照(n=10)粪便 ↑c [38] CRC(n=15);
健康对照(n=12)粪便 ↓e [32] CRC0-Ⅰ期(n=8);
CRCⅡ-Ⅳ期(n=8)血清 ↑f ↑f [39] CRC(n=66);
息肉(n=76);
健康对照(n=92)血清 ↑d↑e ↑d↑e [40] CRC(n=20);
健康对照(n=20)血清 ↑e ↓e [41] CRC(n=20);
健康对照(n=20)粪便 ↓e ↓e ↓e [31] 结直肠腺瘤(n=17);
健康对照(n=17)血浆 ↑c [27] CRC(n=49),
分为非恶病质(n=23);
恶病质前(n=13);
恶病质(n=16)尿液 ↓g [42] 散发性CRC(n=50);
健康对照组(n=50)粪便 ↓e [30] CRC0期(n=30);
CRCI/II期(n=80);
CRCⅢ/IV期(n=68);
多发性息肉状腺瘤(n=45);
健康对照(n=149)粪便 ↓h ↓h ↑c [28] 注:a结直肠癌组织与正常组织相比;b结肠腺瘤和CRC(Ⅲ、Ⅳ期)患者与健康人相比;c息肉或腺瘤患者与健康人相比;d结直肠癌患者与息肉患者相比;e结直肠癌患者与健康人相比;fCRCⅡ-Ⅳ期患者与CRC0-Ⅰ期患者相比;gCRC恶病质前患者与CRC非恶病质患者相比;hCRC 0期患者与健康人相比;↑升高,↓降低。 -
从来源划分,BAs分为初级BAs和次级BAs,由肝细胞中的胆固醇合成的那些称为初级BAs,当初级胆汁酸在肠道中被酶修饰或受细菌作用时,它们变成次级BAs,这种修饰包括核羟基的去除、氧化或差向异构化,包括DCA和LCA等[43]。早期流行病学研究发现,在粪便BAs浓度增加的人群中,结直肠癌的发病率增加[44]。后续研究表明,结直肠癌的发生发展与血清、胆汁和粪便中次级BAs水平的升高有关[45]。Cross等[46]发现,血清中GCDCA的水平与女性结直肠癌风险增加有关,但与男性结直肠癌风险增加无关。LCA也与癌症的发生有关,初步数据表明LCA可杀死神经母细胞瘤细胞和其他可能预防结直肠癌的恶性细胞[47]。DCA和LCA被认为是结肠炎症和结直肠癌的促发因素[48-49],在大肠中作为肿瘤诱导剂[50]。Ridlon等[45]提出,牛磺胆酸(TCA)的微生物代谢可能通过产生硫化氢(基因毒性)和DCA,促进结直肠癌的发生。事实上,在不同的结直肠癌啮齿动物模型中均观察到DCA和LCA在结肠癌变早期阶段作为肿瘤诱导剂。因此,长期暴露于高水平的次级BAs,可能引起炎症和癌症[51]。而高脂肪饮食会增加次级BAs水平[52],被认为是结直肠癌的诱发因素。同样,胆囊切除术因增加了肠道对BAs的暴露,也被认为是结直肠癌的诱发因素。
-
有研究表明[53],高浓度的DCA和LCA均可通过固有的凋亡途径促进细胞凋亡,包括刺激线粒体氧化应激、产生活性氧、细胞色素C(Cytc)释放和激活胞质。另外,许多研究表明,BAs可诱导结肠细胞DNA损伤,例如,在正常结肠细胞系中,LCA可引起DNA损伤,这种损伤会引起复制错误,导致突变,继而造成癌基因和抑癌基因的异常表达[54]。DCA也可以通过引起膜扰动导致花生四烯酸的释放,花生四烯酸被环氧合酶-2酶和脂氧合酶转化为促炎症和促血管生成的前列腺素、活性氧,破坏DNA并抑制DNA修复酶,从而引起DNA损伤[55]。且DCA和7-酮-DCA已被证实与活性氧、活性氮的产生有关,可改变细胞、线粒体膜的稳定性,诱导DNA损伤、突变和凋亡。
BAs在促进肿瘤的发生和进展中的作用已在多个研究中得到了证实,Kitamura等 [56] 发现BAs可直接诱导EGFR和HER2的表达,激活EGFR/HER2通路及其下游通路,从而诱导肿瘤的发生。另外,BAs水平的升高会显著增加MMP7的表达,MMP7高表达的肿瘤具有更强的侵袭性和转移能力[57-58],说明BAs也可能在肿瘤的侵袭和迁移中起作用。有报道称,非结合型BAs作为结肠癌的危险因素,可通过改变毒蕈碱3受体(M3R)和wnt/β-连环蛋白信号通路,诱导结肠上皮发生癌变[59]。也有研究者发现了BAs对肿瘤发生发展的免疫调节机制,Hang等[60]通过对BAs代谢物的筛选,发现了LCA两种不同的代谢产物(3-oxoLCA和isoalloLCA)可调节小鼠T细胞功能,提示BAs代谢物可通过直接调控TH17和Treg细胞平衡,从而调控宿主免疫。类似地,Campbell等[61]研究发现, isoDCA可以促进外周调节性T细胞(P-Treg)生成。上述研究表明3-oxoLCA、isoalloLCA和isoDCA具有潜在的调节肠道免疫细胞的功能,推测可能通过调节肠道免疫系统参与CRC的发生发展。
-
胆汁酸的增加,特别是DCA,与结直肠癌的发展密切相关。相反,UDCA可能具有化学预防特性[62]在一项系统回顾和荟萃分析研究中,Singh等[63]发现,低剂量UDCA可降低PSC-炎症性肠病(IBD)患者晚期CRC的风险。许多体外和体内研究表明,UDCA对结直肠癌有预防作用。有趣的是,Thompson等[64]发现,UDCA仅在男性中可以有效预防结直肠癌。
Brown等[65]研究表明,食用米糠可以抑制结直肠癌小鼠的肿瘤生长,并改变人粪便微生物组。他们发现,结直肠癌患者在摄入米糠4周后,粪便中初级BAs GCDCA和TCA减少了约75%,次级BAs GDCA减少了80%。结直肠癌患者粪便中初级、次级BAs的减少与健康成人一致[66],这被认为是米糠化学预防的一种机制,需要进一步探索。另外,Hanley等[67]提出,甲基供体缺乏可通过影响关键的代谢途径,阻断结直肠癌的进展。与对照组相比,摄入甲基供体缺乏饮食(缺乏叶酸、胆碱、蛋氨酸和维生素B12)的小鼠组织中次级BAs的水平降低,包括DCA、牛磺去氧胆酸(TDCA)、α-MCA、β-MCA和鼠去氧胆酸(MDCA)。因此认为限制甲基供体饮食后,观察到的肿瘤保护作用,可能与次级BAs水平的降低有关。另外,益生菌(双歧杆菌和乳酸杆菌)的摄入可能作为降低DCA水平的手段,从而降低炎症和CRC的风险[68]。因此,通过降低BAs水平,尤其是次级BAs,可能在CRC的预防、治疗中发挥关键作用,但仍需要更加深入的研究。
-
综上所述,BAs根据来源的不同,其作用也大不相同。BAs本身具有抗炎作用;激活后的BAs受体具有免疫调节的作用,也可促进BAs的解毒;在BAs代谢过程中,BAs硫酸化、葡萄糖醛酸化途径是BAs解毒以及清除过多BAs的主要途径,羟基化途径可促进BAs经尿液排泄,推测也与BAs的解毒有关,而酰胺化途径对上皮细胞具有保护作用,被认为是机体的一种防御机制。而次级BAs又具有细胞毒性,可通过诱导细胞凋亡、DNA氧化损伤、细胞增殖增强等机制,诱发炎症和结直肠癌。
许多研究表明,结直肠癌患者存在肠道菌群失调、BAs稳态改变的状况,肠道菌群的失调是导致BAs稳态改变的原因之一,而BAs受体的免疫调节功能可在一定程度上维持BAs稳态,但是否在结直肠癌中起作用尚不明确,有待进一步研究。另外,摄入有益菌、BAs受体激动剂/拮抗剂或直接补充BAs可调节BAs合成,通过减少毒性BAs的产生或清除过多的BAs,可能在结直肠癌的预防、治疗中起作用。进一步阐明BAs在体内的改变及在结直肠癌发生和演变中的作用,明确不同类型BAs同结直肠癌的关联,可能成为结直肠癌预防、诊断和治疗的生物标志物。
Research progress on immunomodulatory effects and role of bile acids and bile acid receptors in the occurrence and development of colorectal cancer
-
摘要: 结直肠癌(CRC)是最常见的恶性肿瘤之一,极大地威胁人类生命健康。胆汁酸稳态的改变可激活相应受体发挥免疫调节功能,同CRC的发生密切相关。此外,部分胆汁酸已被证实可直接诱发结直肠癌,并在结直肠癌的发展过程中发挥重要作用。对胆汁酸的体内代谢过程以及胆汁酸受体的免疫调节作用进行综述,总结了胆汁酸与结直肠癌的关联证据,既往研究显示,调节胆汁酸水平可能起到预防或治疗结直肠癌的作用。Abstract: Colorectal cancer is one of the most common malignant tumors, which is a great threat to human life and health. The change of bile acid homeostasis can activate their corresponding receptors to regulate the immune functions, which is closely related to the occurrence of colorectal cancer. In addition, some bile acids can directly induce colorectal cancer and play an important role in the development of colorectal cancer. In this paper, the metabolic process of bile acids in vivo and the immunomodulatory role of bile acid receptors were reviewed, and the evidence of associations between bile acids and colorectal cancer were summarized, which showed the rebalancing the bile acid levels might play a role in the prevention or treatment of colorectal cancer.
-
Key words:
- colorectal cancer /
- bile acids /
- bile acid receptor /
- immunoregulation
-
图 1 胆汁酸的合成与代谢
注:cholesterol:胆固醇;7α-OH-cholesterol:7α-羟基胆固醇;27-OH-cholesterol:27-羟基胆固醇;25-OH-cholesterol:25-羟胆固醇;24-OH-cholesterol:24-羟基胆固醇;7α-OH-cholesten-3-one(C4):7α-羟基胆甾烯酮;7α,12α-2OH-cholesten-3-one:7α,12α-二羟基胆甾烯酮;CA:胆酸;DCA:脱氧胆酸;G:甘氨酸;T:牛磺酸;CG:甘氨胆酸;TCA:牛磺胆酸;CDCA:鹅去氧胆酸;UDCA:熊去氧胆酸;GCDCA:甘氨鹅去氧胆酸;TCDCA:牛磺鹅脱氧胆酸;LCA:石胆酸;CYP7A1:胆固醇7α-羟化酶;CYP27A1:胆固醇27-羟化酶;Sterol 25-hydroxylase:甾醇25-羟化酶;CYP46A1:CYP450重组酶;HSD3B7:3β-羟基类固醇脱氢酶7;CYP8B1:甾醇-12α-羟化酶;CYP39A1:胆固醇24α7羟化酶;CYP7B1:氧化胆固醇7α-羟化酶;HSDH:3β-羟基类固醇脱氢酶;BSH:胆盐水解酶;7α-DH:7α脱羟酶。
表 1 结直肠癌、息肉或腺瘤患者体内胆汁酸的变化情况
病例/对照 样本 胆汁酸变化情况 文献 CA GCA TCA CDCA GCDCA LCA DCA 7-酮DCA 12-酮DCA 7-OH-3O胆烷酸 CRC组织(n=10);
良性组织(n=9)组织 ↑a ↑a ↑a [34] 结肠腺瘤(n=59);
CRC(n=56,Ⅰ~Ⅳ期各14);
健康对照(n=55)血清 ↓b [36] CRC(n=28);
息肉(n=44);
健康对照(n=55)血清 ↓c ↑d [37] CRC组织(n=17);
邻近黏膜(n=17)组织 ↓a ↓a [32] 结肠腺瘤(n=10);
健康对照(n=10)粪便 ↑c [38] CRC(n=15);
健康对照(n=12)粪便 ↓e [32] CRC0-Ⅰ期(n=8);
CRCⅡ-Ⅳ期(n=8)血清 ↑f ↑f [39] CRC(n=66);
息肉(n=76);
健康对照(n=92)血清 ↑d↑e ↑d↑e [40] CRC(n=20);
健康对照(n=20)血清 ↑e ↓e [41] CRC(n=20);
健康对照(n=20)粪便 ↓e ↓e ↓e [31] 结直肠腺瘤(n=17);
健康对照(n=17)血浆 ↑c [27] CRC(n=49),
分为非恶病质(n=23);
恶病质前(n=13);
恶病质(n=16)尿液 ↓g [42] 散发性CRC(n=50);
健康对照组(n=50)粪便 ↓e [30] CRC0期(n=30);
CRCI/II期(n=80);
CRCⅢ/IV期(n=68);
多发性息肉状腺瘤(n=45);
健康对照(n=149)粪便 ↓h ↓h ↑c [28] 注:a结直肠癌组织与正常组织相比;b结肠腺瘤和CRC(Ⅲ、Ⅳ期)患者与健康人相比;c息肉或腺瘤患者与健康人相比;d结直肠癌患者与息肉患者相比;e结直肠癌患者与健康人相比;fCRCⅡ-Ⅳ期患者与CRC0-Ⅰ期患者相比;gCRC恶病质前患者与CRC非恶病质患者相比;hCRC 0期患者与健康人相比;↑升高,↓降低。  下载: 导出CSV
下载: 导出CSV
-
[1] BRAY F, LAVERSANNE M, SUNG H, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA Cancer J Clin, 2024, 74(3): 229-263. [2] HAN B F, ZHENG R S, ZENG H M, et al. Cancer incidence and mortality in China, 2022[J]. J Natl Cancer Cent, 2024, 4(1): 47-53. [3] LIU Y N, RONG Z H, XIANG D, et al. Detection technologies and metabolic profiling of bile acids: a comprehensive review[J]. Lipids Health Dis, 2018, 17(1): 1-13. doi: 10.1186/s12944-017-0646-8 [4] CHIANG J Y. Bile acid metabolism and signaling[J]. Compr Physiol, 2013, 3(3): 1191-1212. [5] THAKARE R, ALAMOUDI J A, GAUTAM N, et al. Species differences in bile acids : II. bile acid metabolism[J]. J Appl Toxicol, 2018, 38(10): 1336-1352. [6] THAKARE R, ALAMOUDI J A, GAUTAM N, et al. Species differences in bile acids : I. plasma and urine bile acid composition[J]. J Appl Toxicol, 2018, 38(10): 1323-1335. [7] GREIM H, TRÜLZSCH D, CZYGAN P, et al. Mechanism of cholestasis[J]. Gastroenterology, 1972, 63(5): 846-850. doi: 10.1016/S0016-5085(19)33226-3 [8] HIGUCHI H, GORES G J. Ⅳ. Bile acids and death receptors[J]. Am J Physiol Gastrointest Liver Physiol, 2003, 284(5): G734-G738. doi: 10.1152/ajpgi.00491.2002 [9] HUANG J G, BATHENA S P R, CSANAKY I L, et al. Simultaneous characterization of bile acids and their sulfate metabolites in mouse liver, plasma, bile, and urine using LC-MS/MS[J]. J Pharm Biomed Anal, 2011, 55(5): 1111-1119. doi: 10.1016/j.jpba.2011.03.035 [10] TICHO A L, MALHOTRA P, DUDEJA P K, et al. Bile acid receptors and gastrointestinal functions[J]. Liver Res, 2019, 3(1): 31-39. doi: 10.1016/j.livres.2019.01.001 [11] VAVASSORI P, MENCARELLI A, RENGA B, et al. The bile acid receptor FXR is a modulator of intestinal innate immunity[J]. J Immunol, 2009, 183(10): 6251-6261. doi: 10.4049/jimmunol.0803978 [12] TICHO ALEXANDER L, POOJA M, DUDEJA PRADEEP K, et al. Intestinal absorption of bile acids in health and disease[J]. Compr Physiol, 2019, 10(1): 21-56. [13] INAGAKI T, MOSCHETTA A, LEE Y K, et al. Regulation of antibacterial defense in the small intestine by the nuclear bile acid receptor[J]. Proc Natl Acad Sci USA, 2006, 103(10): 3920-3925. doi: 10.1073/pnas.0509592103 [14] D'ALDEBERT E, BIYEYEME BI MVE M J, MERGEY M, et al. Bile salts control the antimicrobial peptide cathelicidin through nuclear receptors in the human biliary epithelium[J]. Gastroenterology, 2009, 136(4): 1435-1443. doi: 10.1053/j.gastro.2008.12.040 [15] GADALETA R M, VAN ERPECUM K J, OLDENBURG B, et al. Farnesoid X receptor activation inhibits inflammation and preserves the intestinal barrier in inflammatory bowel disease[J]. Gut, 2011, 60(4): 463-472. doi: 10.1136/gut.2010.212159 [16] CIPRIANI S, MENCARELLI A, CHINI M G, et al. The bile acid receptor GPBAR-1(TGR5)modulates integrity of intestinal barrier and immune response to experimental colitis[J]. PLoS One, 2011, 6(10): e25637. doi: 10.1371/journal.pone.0025637 [17] HASELOW K, BODE J G, WAMMERS M, et al. Bile acids PKA-dependently induce a switch of the IL-10/IL-12 ratio and reduce proinflammatory capability of human macrophages[J]. J Leukoc Biol, 2013, 94(6): 1253-1264. doi: 10.1189/jlb.0812396 [18] BIAGIOLI M, CARINO A, CIPRIANI S, et al. The bile acid receptor GPBAR1 regulates the M1/M2 phenotype of intestinal macrophages and activation of GPBAR1 rescues mice from murine colitis[J]. J Immunol, 2017, 199(2): 718-733. doi: 10.4049/jimmunol.1700183 [19] ICHIKAWA R, TAKAYAMA T, YONENO K, et al. Bile acids induce monocyte differentiation toward interleukin-12 hypo-producing dendritic cells via a TGR5-dependent pathway[J]. Immunology, 2012, 136(2): 153-162. doi: 10.1111/j.1365-2567.2012.03554.x [20] XIE W, RADOMINSKA-PANDYA A, SHI Y, et al. An essential role for nuclear receptors SXR/PXR in detoxification of cholestatic bile acids[J]. Proc Natl Acad Sci USA, 2001, 98(6): 3375-3380. doi: 10.1073/pnas.051014398 [21] OWEN B M, MILONA A, VAN MIL S, et al. Intestinal detoxification limits the activation of hepatic pregnane X receptor by lithocholic acid[J]. Drug Metab Dispos, 2010, 38(1): 143-149. [22] DE AGUIAR VALLIM T Q, TARLING E J, EDWARDS P A. Pleiotropic roles of bile acids in metabolism[J]. Cell Metab, 2013, 17(5): 657-669. doi: 10.1016/j.cmet.2013.03.013 [23] KIM J H, YAMAORI S, TANABE T, et al. Implication of intestinal VDR deficiency in inflammatory bowel disease[J]. Biochim Biophys Acta, 2013, 1830(1): 2118-2128. doi: 10.1016/j.bbagen.2012.09.020 [24] LIU T Y, SONG X L, KHAN S, et al. The gut microbiota at the intersection of bile acids and intestinal carcinogenesis: an old story, yet mesmerizing[J]. Int J Cancer, 2020, 146(7): 1780-1790. doi: 10.1002/ijc.32563 [25] WU S P, YOON S, ZHANG Y G, et al. Vitamin D receptor pathway is required for probiotic protection in colitis[J]. Am J Physiol Gastrointest Liver Physiol, 2015, 309(5): G341-G349. doi: 10.1152/ajpgi.00105.2015 [26] VAR1ŞCCCL1 B, CAGLAYAN C, KANDEMIR F M, et al. Chrysin mitigates diclofenac-induced hepatotoxicity by modulating oxidative stress, apoptosis, autophagy and endoplasmic reticulum stress in rats[J]. Mol Biol Rep, 2023, 50(1): 433-442. [27] ZHANG Q S, NONG Y Y, LIU Z Q, et al. Proteinase K combining two-step liquid-liquid extraction for plasma untargeted liquid chromatography-mass spectrometry-based metabolomics to discover the potential mechanism of colorectal adenoma[J]. Anal Chem, 2019, 91(22): 14458-14466. [28] YACHIDA S, MIZUTANI S, SHIROMA H, et al. Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer[J]. Nat Med, 2019, 25(6): 968-976. doi: 10.1038/s41591-019-0458-7 [29] BROWN D G, RAO S, WEIR T L, et al. Metabolomics and metabolic pathway networks from human colorectal cancers, adjacent mucosa, and stool[J]. Cancer Metab, 2016, 4: 11. doi: 10.1186/s40170-016-0151-y [30] YANG Y Z, MISRA B B, LIANG L, et al. Integrated microbiome and metabolome analysis reveals a novel interplay between commensal bacteria and metabolites in colorectal cancer[J]. Theranostics, 2019, 9(14): 4101-4114. doi: 10.7150/thno.35186 [31] LE GALL G, GUTTULA K, KELLINGRAY L, et al. Metabolite quantification of faecal extracts from colorectal cancer patients and healthy controls[J]. Oncotarget, 2018, 9(70): 33278-33289. doi: 10.18632/oncotarget.26022 [32] WANG X X, WANG J P, RAO B Q, et al. Gut flora profiling and fecal metabolite composition of colorectal cancer patients and healthy individuals[J]. Exp Ther Med, 2017, 13(6): 2848-2854. doi: 10.3892/etm.2017.4367 [33] BERNSTEIN H, BERNSTEIN C, PAYNE C M, et al. Bile acids as endogenous etiologic agents in gastrointestinal cancer[J]. World J Gastroenterol, 2009, 15(27): 3329-3340. doi: 10.3748/wjg.15.3329 [34] WILLIAMS M D, ZHANG X, PARK J J, et al. Characterizing metabolic changes in human colorectal cancer[J]. Anal Bioanal Chem, 2015, 407(16): 4581-4595. doi: 10.1007/s00216-015-8662-x [35] CHETWYND A J, OGILVIE L A, NZAKIZWANAYO J, et al. The potential of nanoflow liquid chromatography-nano electrospray ionisation-mass spectrometry for global profiling the faecal metabolome[J]. J Chromatogr A, 2019, 1600: 127-136. doi: 10.1016/j.chroma.2019.04.028 [36] UCHIYAMA K, YAGI N, MIZUSHIMA K, et al. Serum metabolomics analysis for early detection of colorectal cancer[J]. J Gastroenterol, 2017, 52(6): 677-694. doi: 10.1007/s00535-016-1261-6 [37] DENG L L, GU H W, ZHU J J, et al. Combining NMR and LC/MS using backward variable elimination: metabolomics analysis of colorectal cancer, polyps, and healthy controls[J]. Anal Chem, 2016, 88(16): 7975-7983. doi: 10.1021/acs.analchem.6b00885 [38] BRIM H, YOOSEPH S, LEE E, et al. A microbiomic analysis in African Americans with colonic lesions reveals Streptococcus sp. VT162 as a marker of neoplastic transformation[J]. Genes, 2017, 8(11): 314. doi: 10.3390/genes8110314 [39] VAHABI F, SADEGHI S, ARJMAND M, et al. Staging of colorectal cancer using serum metabolomics with 1HNMR Spectroscopy[J]. Iran J Basic Med Sci, 2017, 20(7): 835-840. [40] DJUKOVIC D, ZHANG J J, RAFTERY D. Colorectal cancer detection using targeted LC-MS metabolic profiling[M]. Colorectal Cancer. New York: Humana Press, 2018: 229-240. [41] BIAN X Q, LI N, TAN B B, et al. Polarity-tuning derivatization-LC-MS approach for probing global carboxyl-containing metabolites in colorectal cancer[J]. Anal Chem, 2018, 90(19): 11210-11215. doi: 10.1021/acs.analchem.8b01873 [42] OSE J, GIGIC B, LIN T D, et al. Multiplatform urinary metabolomics profiling to discriminate cachectic from non-cachectic colorectal cancer patients: pilot results from the ColoCare study[J]. Metabolites, 2019, 9(9): 178. [43] WAHӏSTOM ӦM A, KOVATCHEVA-DATCHARY P, STAHӏMAN M, et al. Crosstalk between bile acids and gut microbiota and its impact on farnesoid X receptor signalling[J]. Dig Dis, 2017, 35(3): 246-250. [44] LOUIS P, HOLD G L, FLINT H J. The gut microbiota, bacterial metabolites and colorectal cancer[J]. Nat Rev Microbiol, 2014, 12(10): 661-672. doi: 10.1038/nrmicro3344 [45] RIDLON J M, WOLF P G, GASKINS H R. Taurocholic acid metabolism by gut microbes and colon cancer[J]. Gut Microbes, 2016, 7(3): 201-215. doi: 10.1080/19490976.2016.1150414 [46] CROSS AJ, MOORE SC, BOCA S, et al. A prospective study of serum metabolites and colorectal cancer risk[J]. Cancer, 2014, 120(19): 3049-3057. [47] ESTIÚ M C, FRAILUNA M A, OTERO C, et al. Relationship between early onset severe intrahepatic cholestasis of pregnancy and higher risk of meconium-stained fluid[J]. PLoS One, 2017, 12(4): e0176504. doi: 10.1371/journal.pone.0176504 [48] ZENG H W, CLAYCOMBE K J, REINDL K M. Butyrate and deoxycholic acid play common and distinct roles in HCT116 human colon cell proliferation[J]. J Nutr Biochem, 2015, 26(10): 1022-1028. doi: 10.1016/j.jnutbio.2015.04.007 [49] LULU F, PRATIMA N M, EVAN A, et al. Bile acid: a potential inducer of colon cancer stem cells[J]. Stem Cell Res Ther, 2016, 7(1): 181. doi: 10.1186/s13287-016-0439-4 [50] GADALETA R M, GARCIA-IRIGOYEN O, MOSCHETTA A. Bile acids and colon cancer: is FXR the solution of the conundrum?[J]. Mol Aspects Med, 2017, 56: 66-74. doi: 10.1016/j.mam.2017.04.002 [51] VAUGHN B P, KAISER T, STALEY C, et al. A pilot study of fecal bile acid and microbiota profiles in inflammatory bowel disease and primary sclerosing cholangitis[J]. Clin Exp Gastroenterol, 2019, 12: 9-19. doi: 10.2147/CEG.S186097 [52] ZENG H W, UMAR S, RUST B, et al. Secondary bile acids and short chain fatty acids in the colon: a focus on colonic microbiome, cell proliferation, inflammation, and cancer[J]. Int J Mol Sci, 2019, 20(5): 1214. doi: 10.3390/ijms20051214 [53] AMARAL J D, VIANA R J S, RAMALHO R M, et al. Bile acids: regulation of apoptosis by ursodeoxycholic acid[J]. J Lipid Res, 2009, 50(9): 1721-1734. doi: 10.1194/jlr.R900011-JLR200 [54] HARA E. Relationship between obesity, gut microbiome and hepatocellular carcinoma development[J]. Dig Dis, 2015, 33(3): 346-350. doi: 10.1159/000371679 [55] DEBRUYNE P R, BRUYNEEL E A, KARAGUNI I M, et al. Bile acids stimulate invasion and haptotaxis in human colorectal cancer cells through activation of multiple oncogenic signaling pathways[J]. Oncogene, 2002, 21(44): 6740-6750. doi: 10.1038/sj.onc.1205729 [56] KITAMURA T, SRIVASTAVA J, DIGIOVANNI J, et al. Bile acid accelerates erbB2-induced pro-tumorigenic activities in biliary tract cancer[J]. Mol Carcinog, 2015, 54(6): 459-472. doi: 10.1002/mc.22118 [57] FENG H Y, CHEN Y C. Role of bile acids in carcinogenesis of pancreatic cancer: an old topic with new perspective[J]. World J Gastroenterol, 2016, 22(33): 7463-7477. doi: 10.3748/wjg.v22.i33.7463 [58] RAUFMAN J P, DAWSON P A, RAO A, et al. Slc10a2-null mice uncover colon cancer-promoting actions of endogenous fecal bile acids[J]. Carcinogenesis, 2015, 36(10): 1193-1200. doi: 10.1093/carcin/bgv107 [59] LOKE MF, CHUA EG, GAN HM, et al. Metabolomics and 16S rRNA sequencing of human colorectal cancers and adjacent mucosa[J]. PLOS ONE, 2018, 13(12): e0208584. doi: 10.1371/journal.pone.0208584 [60] HANG S Y, PAIK D, YAO L N, et al. Bile acid metabolites control TH17 and Treg cell differentiation[J]. Nature, 2019, 576(7785): 143-148. doi: 10.1038/s41586-019-1785-z [61] CAMPBELL C, MCKENNEY P T, KONSTANTINOVSKY D, et al. Bacterial metabolism of bile acids promotes generation of peripheral regulatory T cells[J]. Nature, 2020, 581(7809): 475-479. doi: 10.1038/s41586-020-2193-0 [62] Centuori S M, Martinez J D. Differential regulation of EGFR-MAPK signaling by deoxycholic acid (DCA) and ursodeoxycholic acid (UDCA) in colon cancer[J]. Dig Dis Sci, 2014, 59(10): 2367-2380. doi: 10.1007/s10620-014-3190-7 [63] SINGH S, KHANNA S, PARDI D S, et al. Effect of ursodeoxycholic acid use on the risk of colorectal neoplasia in patients with primary sclerosing cholangitis and inflammatory bowel disease: a systematic review and meta-analysis[J]. Inflamm Bowel Dis, 2013, 19(8): 1631-1638. doi: 10.1097/MIB.0b013e318286fa61 [64] THOMPSON P A, WERTHEIM B C, ROE D J, et al. Gender modifies the effect of ursodeoxycholic acid in a randomized controlled trial in colorectal adenoma patients[J]. Cancer Prev Res, 2009, 2(12): 1023-1030. doi: 10.1158/1940-6207.CAPR-09-0234 [65] BROWN D G, BORRESEN E C, BROWN R J, et al. Heat-stabilised rice bran consumption by colorectal cancer survivors modulates stool metabolite profiles and metabolic networks: a randomised controlled trial[J]. Br J Nutr, 2017, 117(9): 1244-1256. doi: 10.1017/S0007114517001106 [66] SHEFLIN A M, BORRESEN E C, WDOWIK M J, et al. Pilot dietary intervention with heat-stabilized rice bran modulates stool microbiota and metabolites in healthy adults[J]. Nutrients, 2015, 7(2): 1282-1300. doi: 10.3390/nu7021282 [67] HANLEY M, ALADELOKUN O, KADAVERU K, et al. Methyl donor deficiency blocks colorectal cancer development by affecting key metabolic pathways[J]. Cancer Prev Res, 2019, 13: 1-14. [68] SÁNCHEZ B. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis: a role for bifidobacteria and lactobacilli?[J]. Nat Rev Gastroenterol Hepatol, 2018, 15(4): 205. -

 点击查看大图
点击查看大图
图(1) / 表(1)
计量
- 文章访问数: 10275
- HTML全文浏览量: 4840
- PDF下载量: 41
- 被引次数: 0
