

 下载:
下载:



-
细胞间隙连接(Intercellular gap junction, GJIC)是一种存在于人体所有细胞中的膜通道,由连接蛋白(connexins, Cxs)形成,并负责转移生物活性分子、代谢物和相邻细胞或细胞与细胞外环境间的盐离子,对细胞的增殖、分化及机体内环境稳定、新陈代谢、生长发育起至关重要作用[1]。实验证实小鼠骨髓、肝脏及脾脏基质中有11种不同的连接蛋白表达,但人类骨髓基质中仅仅有3种Cxs (Cx31、Cx43、Cx45)表达。多项实验均证实Cx43在支持正常造血过程中具有重要作用,而我们前期的研究发现,Cx43在多发性骨髓瘤(multiple myeloma,MM)的发病过程中具有重要作用,患者骨髓微环境中的Cx43表达水平较正常明显升高,骨髓瘤细胞与成骨细胞相互作用后可通过由Cx43组成的GJIC促进其迁移,上调Cx43表达对多发性骨髓瘤细胞的增殖及迁移均起到促进作用,Cx43表达异常与骨髓瘤融合细胞发生相关[2-3]。然而Cx43在MM细胞生存及耐药中的作用尚未阐明,尤其在多发性骨髓瘤干细胞及其与微环境中作用尚不明确。有鉴于此,本研究分离、培养MM患者及正常志愿者来源骨髓间充质干细胞(MM-MSCs、ND-MSCs),在直接共培养条件下观察MM干细胞样细胞生物学特性的变化及MM-MSCs对MM干细胞样细胞的生存及耐药的作用,并探讨其可能机制。
-
细胞株:MM细胞株RPMI 8226、U266、XG4、XG7(苏州大学生物技术研究所张学光教授惠赠);试剂:FBS、PBS、LG-DMEM完全培养液、RPMI1640培养基(美国Gibco公司);Midi MACs系统(德国Milteyni公司);Hoechst33342(美国Sigma公司);抗Cx43及GAPDH一抗(美国CST公司);RNeasy kit试剂盒、QuantiTect reverse transcriptase kit试剂盒、TopTaq Master Mix Kit 试剂盒(美国Qiagen公司);Cytometric Beads Array试剂盒(美国BD公司)。
-
参考文献[4]的方法,采用Ficoll分离MM患者及正常志愿者骨髓单个核细胞(BM-MNCs),用含10%FBS的LG-DMEM完全培养液培养,观察细胞状态,72 h后首次换液,以后根据情况每2~3 d,换液1次。待细胞生长至80%融合后,胰酶消化传代。传至第三代后收获细胞进行后续实验,剩余细胞标记后冻存于液氮罐中备用。志愿者及患者骨髓间充质干细胞(BM-MSCs)的分离、扩增和鉴定在知情同意下获得,并经医院伦理委员会批准。
-
RPMI 8226、U266采用含有10% FBS的RPMI1640培养基培养。XG4、XG7采用含有10% FBS、1 ng/ml IL-6的RPMI1640培养基培养。原代MM细胞来自6例初诊MM患者骨髓:用Ficoll分离BM-MNCs,并用Midi MACs系统纯化,留取CD38+、CD138+细胞,操作按说明书进行,分选后的细胞采用流式细胞术(FCM)检测其纯度,CD38+、CD138+细胞≥90%,采用含有10% FBS的RPMI1640培养基培养。观察细胞状态,48 h后首次换液,以后根据情况每1~2 d,换液1次。传至第三代后收获细胞进行后续实验,剩余细胞标记后冻存于液氮罐中备用。
-
取对数生长期F3代MM-MSCs及ND-MSCs,用PBS洗涤后,调整细胞浓度为2.0×106/ml,每取100 μl细胞悬液,分别加入PE标记的CD90、CD73、CD44、CD105、CD34、CD45及HLA-DR,阴性对照为PE标记的同型IgG,室温下孵育30 min,PBS洗涤2次后,FCM上机检测。
-
按文献[5]报道的方法,分别取RPMI 8226、U266、XG4、XG7及原代MM细胞,调整细胞浓度为106/ml,加入浓度为1 mg/ml的Hoechst33342,调整其终浓度为5 μg/ml,混匀后置于37 ℃水浴箱中避光孵育120 min,期间数次晃动离心管。对照组于此步骤中加入终浓度为50 μmol/L维拉帕米同时孵育。离心后PBS洗涤,用含碘化丙啶(2 μg/m1)的4 ℃预冷PBS重悬细胞,并置于冰浴中。FCM上机检测,激发波长为350 nm,采集波长为450 nm(蓝光)和675 nm(红光),通过与对照组比较,选取染色偏弱部分的细胞即为SP细胞。SP细胞分选按上述步骤准备细胞,ALTRA流式细胞仪更换鞘液并用酒精进行清洗后换为双蒸水冲洗;分别上Hoechst33342管和Hoechst33342+verapamil管进行检测,FCM选择SP分选方案,调整分选参数,全程需要振荡,分选结束后在无菌条件下分别收集主群细胞(MP)和侧群细胞(SP),备用。
-
分别收集RPMI8226、SP细胞、ND-MSCs、MM-MSCs、SP细胞+ND-MSCs、SP细胞+ND-MSCs+25 mmol/L α-GA、SP细胞+MM-MSCs和SP细胞+MM-MSCs+25 mmol/L α-GA各组细胞,用预冷的PBS洗涤细胞3次,加入细胞裂解液,置4 ℃作用30 min,12000 g/min离心10 min,收集上清液,BCA法测定蛋白浓度,加入4×SDS凝胶加样缓冲液混匀,煮沸10 min使蛋白变性。然后,行聚丙烯酰胺凝胶电泳(SDS-PAGE),并转移至PVDF膜上,封闭1 h后,分别与抗Cx43及GAPDH一抗4 ℃孵育过夜,TBS液洗涤后再与HRP标记的二抗共孵育1 h,洗涤后,应用ECL化学发光法显象和Image图象分析软件分析。
-
采用碘化吡啶(PI)法。实验分组:①SP细胞+MM-MSCs;②SP细胞+MM-MSCs+25 mmol/L 18α甘草次酸(α-GA);③对照组为RPMI 8226细胞。实验设3复孔,FCM分析其DNA含量,CellQuest软件分析结果。
-
采用甲基纤维素半固体培养法。实验分组:①SP细胞+MM-MSC组;②SP细胞+MM-MSC+25 mmol/L α-GA组。分别调整SP细胞和MM-MSC细胞浓度为4×105/ml和2.0×106/ml,与等量的2%甲基纤维素混均后,接种于6孔板,每孔总体系2 ml;置饱和湿度、37 ℃的CO2培养箱中培养,14 d取出,置倒置显微镜下记录集落数,≥50细胞为集落,≤50则为簇。
-
采用逆转录聚合酶链式反应(RT-PCR)方法。实验分组:①RPMI 8226组;②新鲜分离SP细胞组;③SP细胞+MM-MSC组;④SP细胞+MM-MSC+25 mmol/L α-GA组。收集各组细胞,操作按试剂盒说明进行。简述如下:采用RNeasy kit试剂盒提取RNA样本,取1μg RNA进行逆转录,按等量cDNA进行PCR反应。所有引物序列均由上海生物工程公司设计并合成,采用β-actin为内参。β-actin上游引物 5′-TCCTGTGGCATCCACG AAACT-3′,下游引物 5′-GAAGCATTTGC GGTGGACGAT-3′,其它引物见表1。PCR扩增条件均为:94 ℃ 5 min、94 ℃ 40 s、56 ℃ 30 s、72 ℃ 32 s,共35个循环。取4 μl PCR产物、Marker 3.5 μl分别加样于2.0%琼脂糖凝胶中电泳,电压100V电泳30~60 min,紫外投射仪观察目标条带,摄影,图象分析软件Smartview2001分析处理结果。
表 1 干细胞样基因引物序列
基因 引物序列 c-myc 5′CTTCTCTCCGTCCTCGGATTCT
3′GAAGGTGATCCAGACTCTGACCTTKlf-4 5′GCAAGTCCCCTCTCTCCATTA
3′GTAAGGTTTCTCGCCTGTGTGOct-4 5′GGAGATATGCAAAGCAGAAACC
3′CTCAAAATCCTCTCGTTGTGCSox-2 5′CGGCAACCAGAAAAACAGC
3′TCTCCGTCTCCGACAAAAGT -
采用CBA检测法。取对数生长期MM-MSCs,调整细胞数1×105/ml 接种于6孔板,培养箱静置4 h弃上清,并将不同MM细胞按1×105/ml的浓度接种该孔中,每孔2 ml,分组为:①RPMI 8226细胞;②SP细胞+MM-MSCs;③SP+MM-MSCs+α-GA(25mmol/L);④MM-MSCs。每组设3个复孔,培养24 h后收集培养上清,利用CBA技术测定上清中IL-6、IL-10、TGFβ、bFGF和IL-17的变化。
-
采用annexinV/PI标记细胞流式术分析法。取对数生长期MM-MSC细胞,调整细胞数4×105/孔接种于24孔板;RPMI8226或SP细胞,调整细胞数2×104/孔接种于24孔板,培养箱静置4 h后去上清,分组如下:①RPMI8226;②RPMI8226+硼替佐米(BTZ);③SP+BTZ;④SP+MM-MSC+BTZ;⑤SP+MM-MSC+BTZ+α-GA。所有实验组BTZ及α-GA的终浓度分别为20 nmol/L和25 mmol/L,培养24 h后收集细胞,FCM检测细胞凋亡,实验设5复孔。
-
所有数据采用Graphpad Prism 5.0 统计处理软件分析,以均数±标准差表示。组间分析采用t检验,P<0.05 为差别具有统计学意义。
-
分离培养获得MM-MSCs及ND-MSCs,表面抗原提示两者均为高表达CD73(98.0%)、CD44(100%)、CD90(99.8%)和CD105(100%),基本不表达CD34(0.3%)、HLA-DR(0.2%),细胞形态两者无明显差异。见表2和图1。

图 1 BM-MSCs细胞表面抗原表达R
表 2 BM-MSCs细胞表面抗原分析
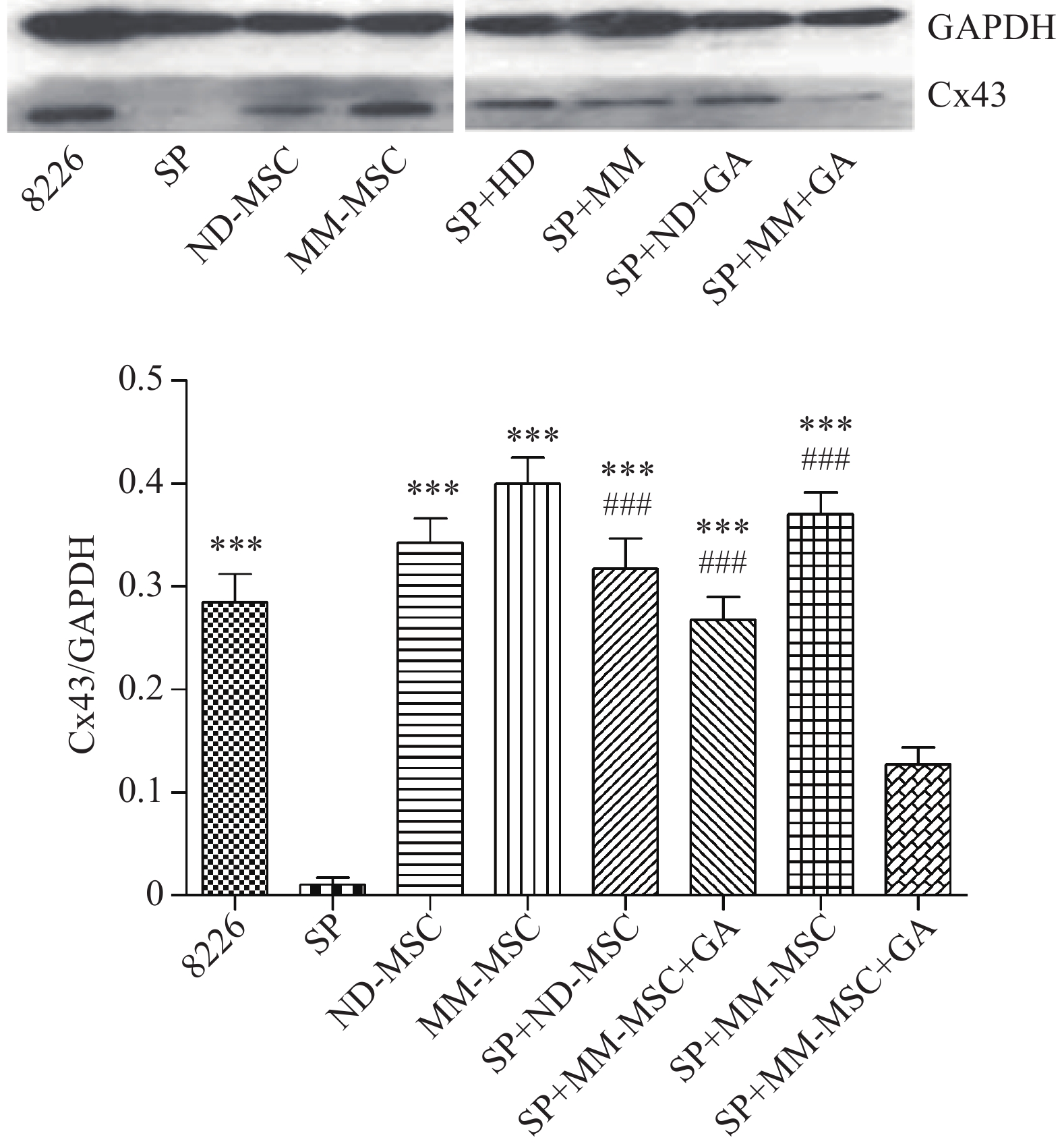
表面抗原 CD73 CD44 CD90 CD105 CD34 HLA-DR 表达率(%) 98.00 100 99.80 100 0.30 0.20 蛋白印迹试验证实SP细胞仅表达极少量的Cx43分子,而RPMI 8226细胞则表达较高水平的Cx43,两者具有显著性差异(P<0.001);MM-MSCs较ND-MSCs表达Cx43明显较多,但不具有统计学意义(P>0.05);SP细胞与MM-MSCs共培养后,其Cx43表达均有显著上调(P<0.001);阻断GJ后,SP细胞的Cx43表达则呈现明显下调(P<0.001),详见图2。

图 2 蛋白印迹分析不同细胞Cx43的表达
-
本研究对6例MM患者的原代细胞及4种MM细胞株的检测提示,采用Hoechst 33342染色后应用FCM技术可将MM细胞分为2群,即主群细胞(MP)和侧群细胞(SP)[6]。所有MM细胞均存在不同比例的SP细胞。MM细胞株中SP细胞含量分别为1.783 %、0.8256 %、0.082 %、0.177 %,而原代细胞不具备可重复性,鉴于RPMI 8266细胞中SP细胞含量较多,且稳定,此后实验采用的SP细胞均来自RPMI8226,详见图3。

图 3 不同MM细胞株的SP流式术分选
-
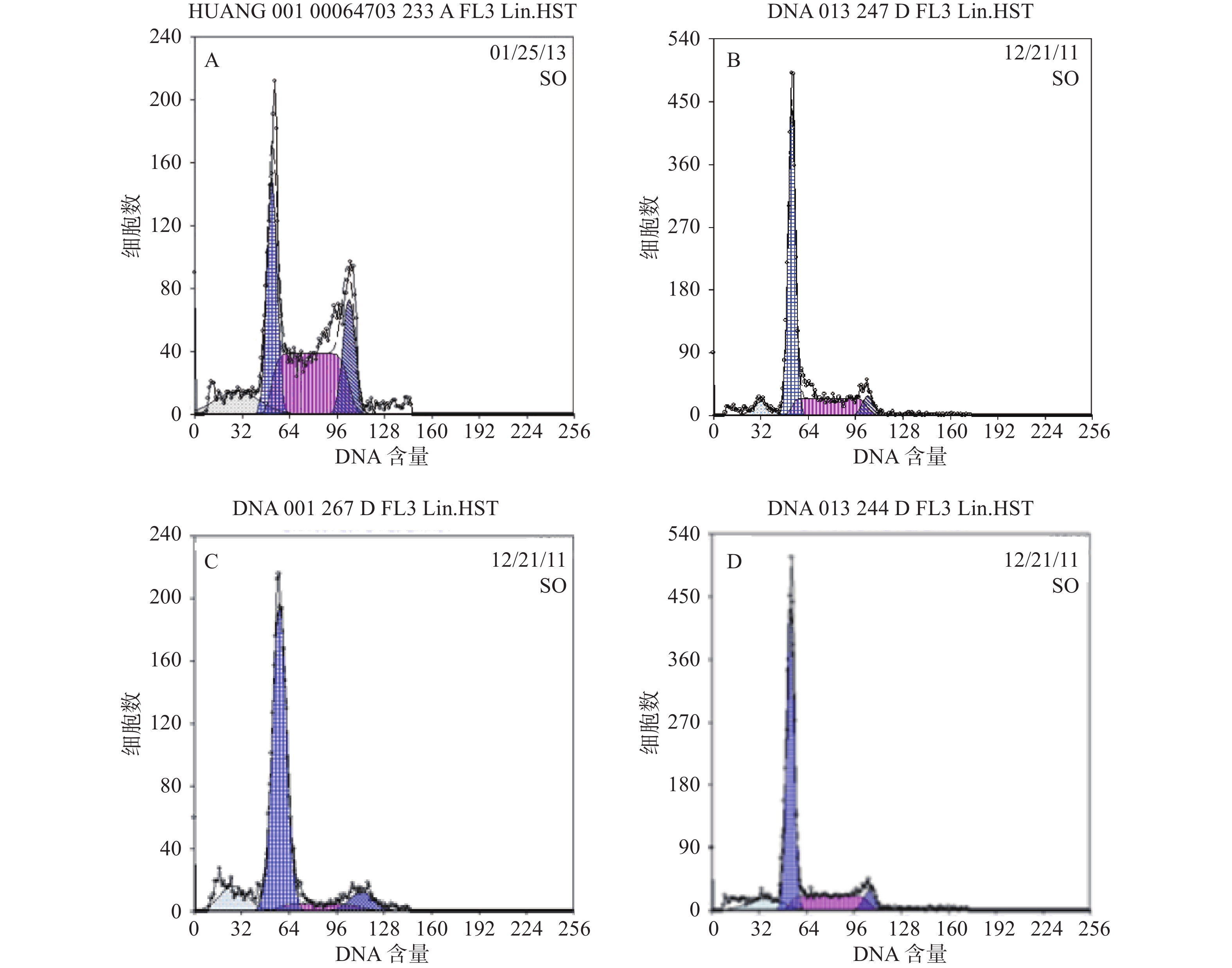
结果分析提示SP亚群中处G0期细胞比例显著高于MP亚群,分别为(44.34±1.7) %和(28.49±1.1) % ,提示SP亚群中包含更多处静止期的MM细胞。与MM-MSCs共培养后发现MM-MSCs具有促进SP亚群细胞进入G0期的作用,其G0期细胞达(82.6±0.1) % (P<0.001),而加入间隙连接抑制剂α-GA后,MM-MSCs对SP亚群的这一作用减弱,细胞进入增殖周期者增多,G0期细胞降至(63.42±3.86) % (P<0.01),详见图4。

图 4 MM-MSCs对SP细胞周期的作用
-
我们利用克隆形成实验分析SP细胞体外形成集落的能力,SP细胞单独培养、与ND-MSCs共培养、与MM-MSCs共培养、与ND-MSCs共培养体系中加入通道阻断剂,与MM-MSCs共培养体系中加入通道阻断剂后单克隆直径、克隆形成数、克隆形成率见表3。结果显示出与MM-MSCs共培养的SP细胞有更强的克隆形成能力。加入通道阻断剂后克隆形成能力均表现出一定程度的下降,单细胞克隆直径减小,克隆形成率降低,见图5。
表 3 不同培养体系加入阻断剂前后克隆形成能力
组别 单克隆直径
(cm)克隆形成数 克隆形成率
(/2000)SP 0.28±0.16 1722±127 86%±6% SP+ND-MSCs 0.33±0.14 1858±89 93%±4% SP+MM-MSCs 0.38±0.21 1900±85 95%±4% SP+ND-MSCs+GA 0.25±0.22 1532±112 77%±6% SP+MM-MSCs+GA 0.31±0.17 1755±76 88%±4% 
图 5 ND-NSCs及MM-MSCs对SP细胞的体外集落形成的影响
-
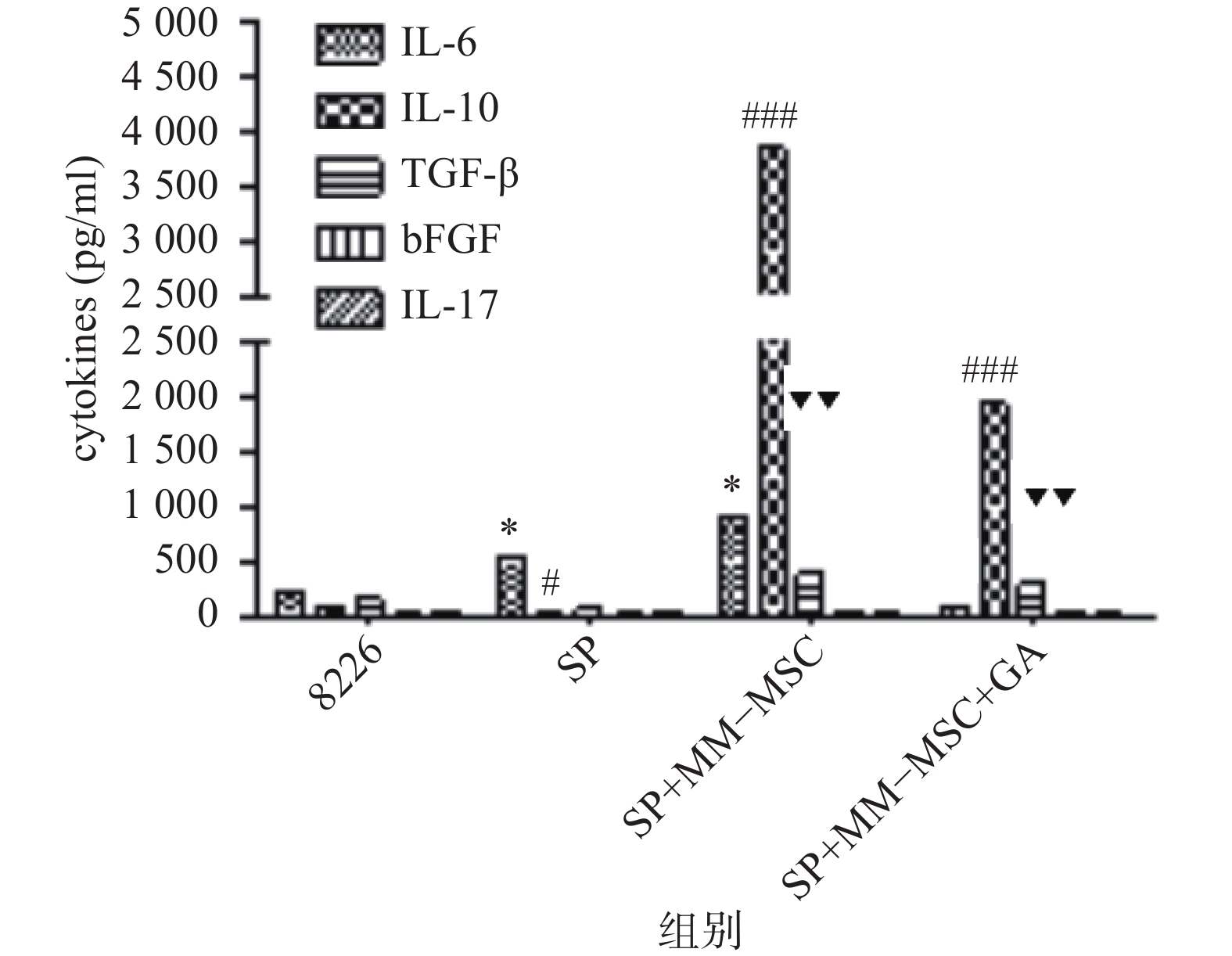
CBA分析显示,MM-MSCs单独培养24 h后,其培养上清中存在高水平的IL-6,较低水平的TGF-β、bFGF和IL-17,基本无IL-10分泌;RPMI 8266细胞培养24 h后上清中可以测得较低水平的TGF-β及少量bFGF、IL-17、IL-6及IL-10;共培养24 h后,其上清中IL-6、IL-10和TGF-β水平较前明显升高(P<0.05),尤其是IL-6和IL-10水平较单独培养时显著升高(P<0.01),bFGF和IL-17共培养前后则无明显变化;加入GJ阻断剂后,细胞因子IL-6、IL-10和TGF-β的分泌有所降低(P<0.05),见图6。

图 6 培养上清中细胞因子变化
-
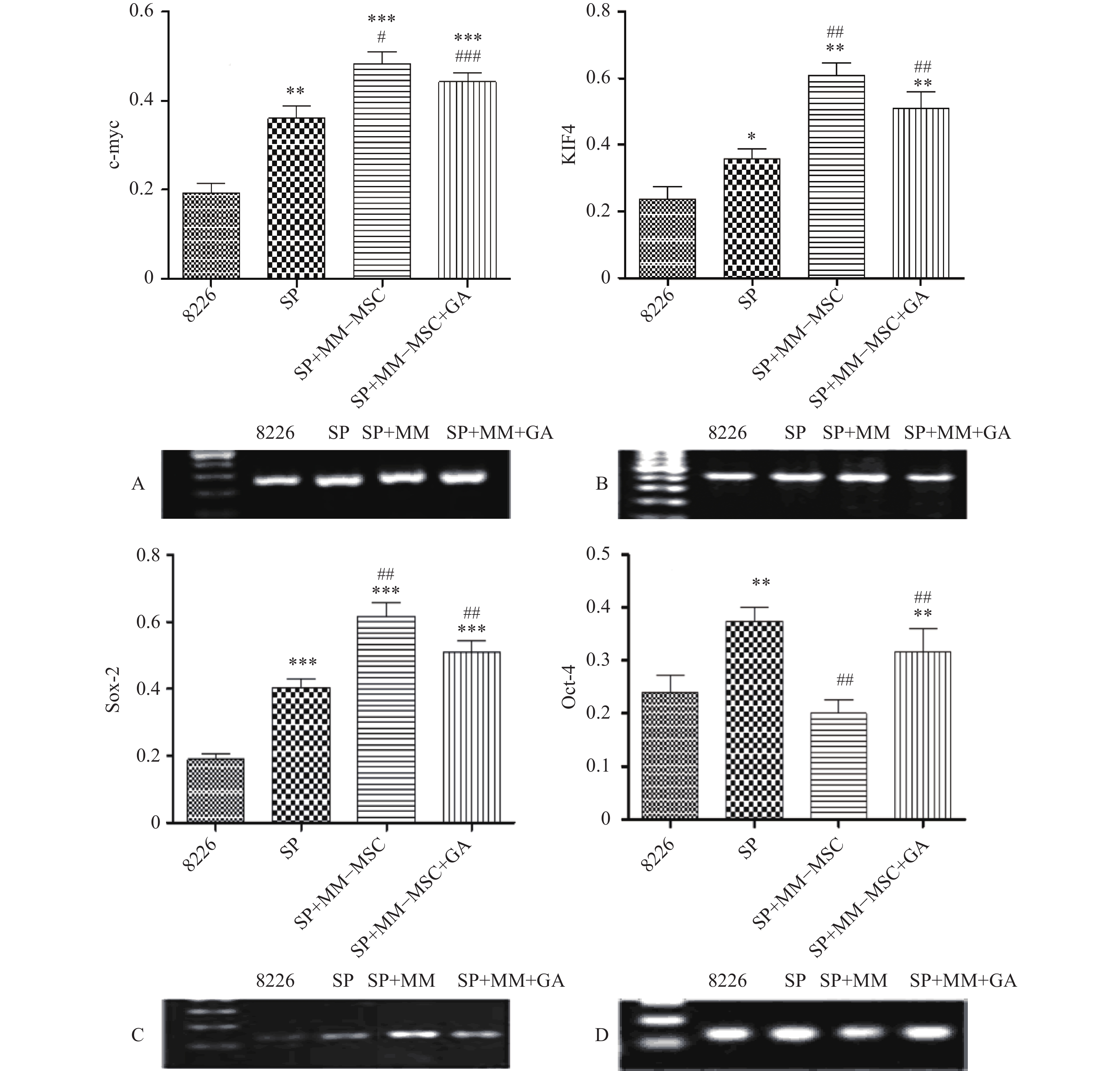
RT-PCR检测发现RPMI8266存在一定量c-myc、KIF4、SOX2和Oct-4基因表达,但SP细胞亚群中该类基因表达明显上调,两者具有显著性差异(P<0.05),将SP细胞与MM-MSC共培养后,可观察到c-myc、KIF4和SOX2基因表达的显著上调(P<0.001),而Oct-4基因表达下调,加入GJ阻断剂后,原上调的基因均有不同程度下调,但无明显区别(P>0.05),见图7。

图 7 RT-PCR分析SP细胞干细胞相关基因表达
-
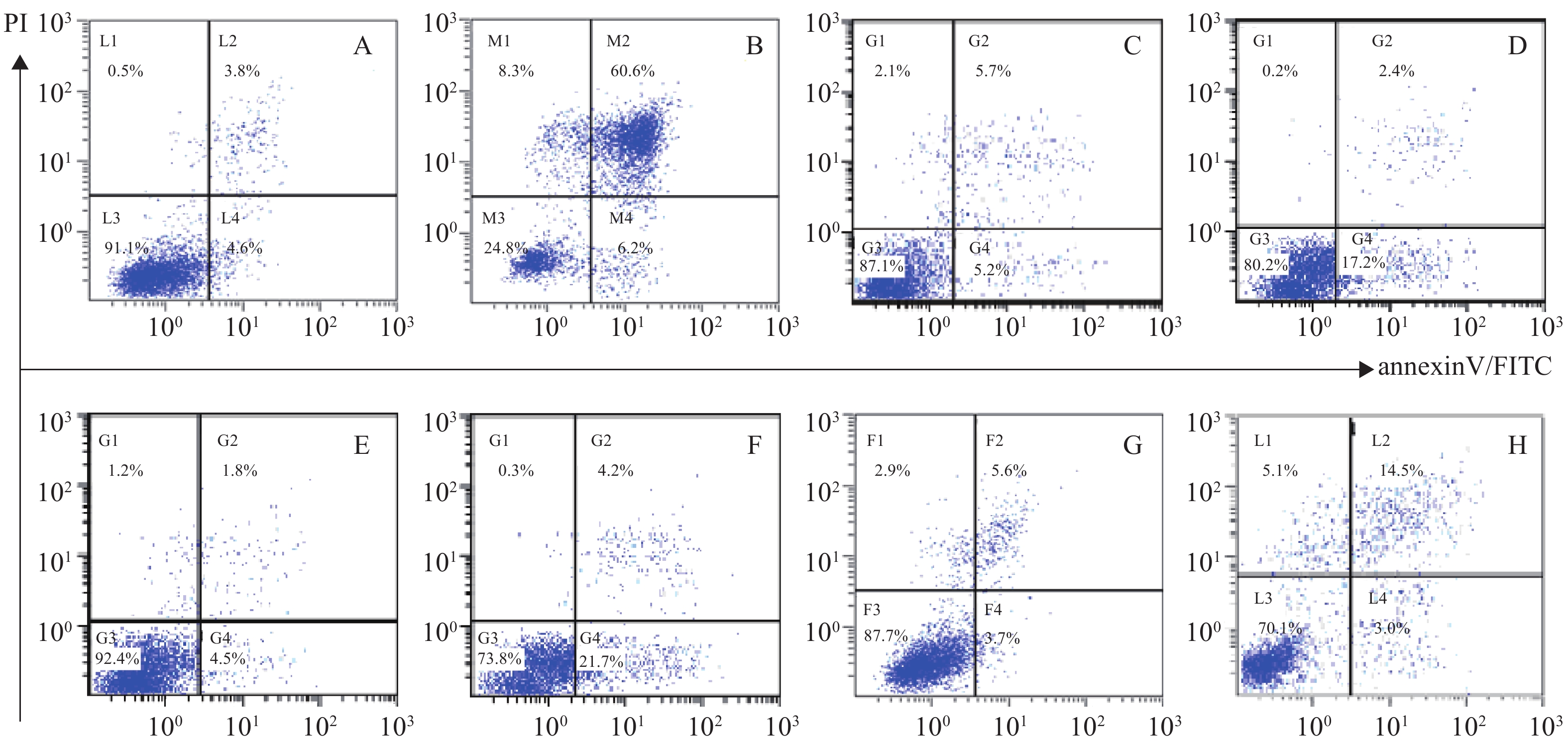
体外PI/Annexin V检测显示,RPMI 8226的MP细胞对BTZ诱导的细胞凋亡敏感,而对SP细胞敏感性较差,其凋亡率分别为(66.8±0.77)%和(25.9±0.86)%,P<0.001。与MM-MSCs直接共培养后,BTZ诱导的凋亡作用较单独培养明显减弱(P<0.05),MM-MSCs具有一定保护作用,加入GJ阻断剂后,可部分恢复MM细胞对硼替佐米的敏感性,证实MM-MSCs可保护骨髓瘤细胞免受抗肿瘤药物影响,而GJIC在其中可能起到一定作用,见图8。

图 8 GJ在硼替佐咪诱导的MM细胞凋亡中的作用
-
MM是一种恶性浆细胞疾病,以肿瘤细胞与骨髓微环境中基质细胞的复杂的相互作用网络为特征,骨髓基质细胞可促进MM细胞的生存、增殖和药物抗性。骨髓间充质干细胞是骨髓基质细胞中最主要的干细胞群体,能够分化成多种细胞系,包括成纤维细胞、脂肪细胞、软骨细胞和成骨细胞。MSCs可以迁移到原发肿瘤和转移部位,这意味着这些细胞可能调节肿瘤的生长和转移。MSCs在与MM细胞相互黏附及作用过程中部分细胞特性发生改变,成为肿瘤相关BMSCs,即MM-MSCs,MM患者来源的骨髓间充质干细胞显示出功能异常,说明骨髓间充质干细胞在骨髓瘤的发生、发展中不是旁观者,它还可分泌多种细胞因子影响MM细胞的生长、生存、耐药、迁移,在疾病的发生、发展中起了重要的作用,目前成为治疗多发性骨髓瘤的新的研究热点[7-8]。
尽管近几年MM的治疗取得了长足的进步,但目前它仍然是不可治愈的,有证据表明MM细胞之间存在异质性,尤其是可能存在MM干细胞亚群,它具有自我更新及原发耐药特性,可能是MM增殖、维持MM表型及导致疾病复发的原因,但其耐药性的机制还没有得到充分的了解[5, 9]。理想的多发性骨髓瘤干细胞(MMSCs)的鉴定应依赖于MMSCs的表型,但至今MMSCs的表型尚未得到正确的定义。Goodell等通过FCM在MM细胞中分离出具有干细胞特性的SP细胞,为拓展肿瘤干细胞的研究提供了思路。目前,SP细胞和ALDH1+已经被用来鉴定MMSCs。MMSCs与骨髓微环境的复杂的相互作用维持着MMSCs的自我更新和生存。然而, MMSCs与周围骨髓微环境相互作用的分子需要进一步确认。
在过去的几年里,连接蛋白的异常表达,特别是Cx43的异常表达已经被证实与癌症复发、转移性扩散和不良预后相关。根据癌症的不同分期和类型,Cx43既可以作为肿瘤抑制因子,也可以作为癌基因、生物标记物,我们需要更好地了解Cx43在肿瘤微环境中如何参与、影响肿瘤形成和进展,从而开发基于Cx43的临床可行疗法。Cx43分子在细胞表面形成通道,可使小分子和一定量大分子物质在细胞间转移,这一特性使它们存在将化疗药物直接送入肿瘤细胞内的潜力,从而成为肿瘤治疗新的非常有吸引力的靶点。
为明确MM-MSCs与SP细胞的相互作用,首先我们采用Hoechst33342标记FCM术检测骨髓瘤细胞株及新鲜MM标本,结果证实所有检测样本中均存在SP细胞亚群,4 种 MM 细胞株检测SP 细胞含量分别为1.783 %、0.8256 %、0.082 %、0.177 %,并成功分选SP细胞比例最高的PRMI8226细胞株的SP细胞。在此基础上,通过对SP亚群细胞的多种干细胞相关基因如c-myc、KIF4、SOX2和Oct-4表达分析,发现其与RPMI 8226细胞有明显不同。直接与MM-MSCs共培养后SP细胞亚群c-myc、KIF4和SOX2基因表达显著上调 (P<0.001),而Oct-4基因表达下调,加入GJIC阻断剂后,c-myc、KIF4和SOX2基因表达尽管有所下调,但并无显著性差异。对于细胞周期的分析也证实SP细胞亚群中处G0期细胞比例显著高于MP亚群,分别为(44.34±1.7)%和(28.49±1.1)% ,提示SP细胞亚群中包含较多处静止期MM细胞,推测与该群细胞化疗敏感性差可能有关。共培养后发现MM-MSCs有促进SP亚群细胞进入G0期的作用(P<0.001),其G0期细胞达(82.6±0.1)%,而加入间隙连接抑制剂α-GA后,这一作用减弱,细胞进入周期者增多,G0期细胞为(63.42±3.86)%。体外集落形成试验证实从MM细胞分离的SP细胞本身具有较强的集落形成能力,但在MM-MSCs存在的前提下其细胞形成的体外集落细胞数更多,胞体较大且细胞折光性强,提示细胞活力较好,培养体系中加入GJ阻断剂后体外集落形成能力均现出一定程度的下降,克隆形成率降低,单细胞克隆直径减小。
前期我们的研究证实MM-MSCs与MM细胞可形成功能性GJIC,并在多发性骨髓瘤的发病中扮演重要角色[2-3, 10],为进一步观察间充质干细胞中连接蛋白Cx43组成的细胞间隙连接在多发性骨髓瘤SP细胞生存及耐药中的作用,我们通过蛋白印迹试验证实除SP细胞基本无Cx43表达外,ND-MSCs和MM-MSCs、MM细胞株及初诊患者MM细胞均表达Cx43分子,与SP细胞相比差别显著(P<0.001)。SP细胞与MM-MSCs共培养后,其Cx43分子表达显著上调(P<0.001),加入α-GA可部分下调SP细胞的Cx43表达(P<0.001)。采用CBA技术检测共培养前后培养液中细胞因子变化,发现在直接共培养时MM-MSCs细胞因子分泌谱发生变化,在原有高水平IL-6的基础上,IL-10和TGFβ水平较前明显增加(P<0.05),尤其是IL-6和IL-10较单独培养时显著增加(P<0.01),bFGF和IL-17在共培养前后水平无明显变化,采用α-GA阻断MM-MSCs与SP细胞间的GJIC后,细胞因子IL-6、IL-10和TGF-β的分泌能力下调(P<0.05)。综上,我们认为MM-MSCs通过多种途径影响SP细胞的生物学特性,增加其干细胞相关基因表达、增加静止期细胞比例、增强其体外集落形成能力、改变细胞因子分泌,而在这一过程中Cx43分子的表达及其形成的GJIC具有重要作用。
蛋白酶体抑制剂BTZ是MM治疗的一线药物,尽管BTZ较目前其他化疗药物更为有效,但原发性或获得性耐药仍是限制其疗效的主要原因,目前对于蛋白酶体抑制剂耐药的机制仍未阐明。本研究发现尽管MP和SP细胞均对BTZ敏感,但SP细胞敏感性较差,可能与其本身极低表达Cx43部分相关,其凋亡细胞分别为(66.8±0.77)%和(25.9±0.86)%,P<0.001,经与MM-MSCs直接共培养后,BTZ诱导的凋亡作用明显减弱(P<0.05),MM-MSCs具有保护作用,而在共培养体系中加入α-GA,可部分恢复MM细胞对硼替佐咪的敏感性,由此证实MM-MSCs促进MM SP细胞的生存,保护MM SP细胞免受抗肿瘤药物影响,GJIC在其中起到一定作用。我们前期实验提示,过表达MM细胞的Cx43可提高MM细胞化疗敏感性,但与MSCs共培养后敏感性降低,肿瘤细胞本身Cx43半通道有可能通过增加癌细胞对化疗药物的通透性来降低耐药性,但与微环境中MSCs相互作用后这一作用产生变化,但具体机制仍在进一步研究中。
本研究发现Cx43及其组成的GJIC不但影响MM细胞的迁移,而且也影响恶性浆细胞的药物敏感性,证实骨髓微环境中Cx43及其组成的GJIC在MM发生、发展中具有重要作用,在细胞因子分泌、肿瘤生长、细胞周期变化、基因表达等多个环节中影响MM细胞的生物学行为,基于我们及其他相关研究结果,我们证实MM细胞与MM-MSCs、造血细胞及细胞外基质通过粘附GJIC介导耐药,阻断肿瘤细胞与微环境的GJIC将有助于提高抗肿瘤效益。
Role of intercellular junctions in the biological behavior of SP cells of multiple myeloma
-
摘要:
目的 观察不同来源间充质干细胞(MSCs)中由连接蛋白43(Cx43)组成的细胞间隙连接(GJIC)及其介导的信号对多发性骨髓瘤(MM)侧群细胞(SP细胞)生物学行为的影响,并探讨其可能机制。 方法 分离培养不同来源的间充质干细胞(MSCs);应用流式细胞术分选MM细胞株RPMI 8266的SP细胞;采用RT-PCR技术及蛋白印迹(Western blot)法检测不同来源MSCs、RPMI 8266、SP细胞中Cx43基因及蛋白水平表达;直接共培养观察不同来源MSCs对SP细胞周期、Cx43蛋白表达、体外集落形成能力、干细胞相关基因表达、细胞因子分泌和耐药的变化以及加入连接通道抑制剂18α甘草次酸(α-GA)后的影响。 结果 MM-MSCs与ND-MSCs形态及表型无明显区别,与RPMI 8266细胞均表达较高水平的Cx43;与MM-MSCs共培养可使更多SP细胞进入G0期(P<0.001),SP细胞的c-myc、KIF4和SOX2基因表达显著上调,而Oct-4基因表达下调,加入α-GA后,c-myc、KIF4和SOX2均有不同程度下调,但无显著性差别;使Cx43表达上调,分别为(31.00±2)%和(39.00±2)%;使体外集落形成能力上调,加入α-GA可部分抑制该作用;RPMI 8266存在少量c-myc、KIF4、SOX2和Oct-4基因表达,SP细胞亚群中该类基因明显上调,MM-MSCs分泌高水平的白介素(IL)-6,与SP细胞共培养后,其上清液中IL-6、IL-10及TGF-β表达上调(P=0.0072,P=0.037);bFGF和IL-17则无明显变化。加入α-GA后,上清液中IL-6、IL-10和TGF-β水平降低;MM细胞对硼替佐米诱导的凋亡敏感,但SP细胞敏感性较差,与MM-MSCs共培养显著减少硼替佐米介导的细胞凋亡,加入α-GA可部分恢复MM细胞对硼替佐米的敏感性。 结论 MM-MSCs与多发性骨髓瘤SP细胞上调Cx43蛋白表达,形成更多GJIC,并通过改变MSCs细胞因子分泌谱,促进SP细胞增殖和耐药,可能是最终导致MM复发的原因之一。 Abstract:Objective To observe the effects of the intercellular gap junction (GJIC) composed of connexin 43(Cx43) in mesenchymal stem cells (MSCs) from different sources and their signals on the biological behavior of multiple myeloma (MM) lateral population cells (SP cells), and to explore its possible mechanism. Methods Mesenchymal stem cells (MSCs) from different sources were isolated and cultured. SP cells of MM cell line RPMI 8266 were sorted by flow cytometry. RT-PCR and Western blot were used to detect the expression of Cx43 gene and protein in MSCs, RPMI 8266 and SP cells from different sources. The effects of MSCs from different sources on SP cell cycle, Cx43 protein expression, colony formation ability in vitro, stem cell related gene expression, cytokine secretion and drug resistance were observed. Results There was no significant difference in morphology and phenotype between MM-MSCs and ND-MSCs. Both MM-MSCs and RPMI 8266 cells expressed a higher level of Cx43. Co-culture with MM-MSCs induced more SP cells to enter G0 phase (P<0.001). The expressions of c-myc, Kif4 and Sox2 genes in SP cells were significantly up-regulated, while the expression of Oct-4 gene was down-regulated. After adding α-GA, c-myc, Kif4 and Sox2 were down-regulated in varying degrees, but there was no significant difference. The expression of Cx43 was up-regulated by (31.00±2)% and (39.00±2)%, respectively. The colony formation ability in vitro was up-regulated, and the addition of α-GA could partially inhibit this effect. A small amount of c-myc, Kif4, Sox2 and Oct-4 genes were expressed in RPMI 8266. These genes were significantly up-regulated in SP cell subpopulation. MM-MSCs secreted high levels of interleukin (IL)-6. After co-culture with SP cells, the expressions of IL-6, IL-10 and TGF-β in the supernatant of MM-MSCs were up-regulated (P=0.0072, P=0.037). bFGF and IL-17 had no significant change. After adding α-GA, the levels of IL-6, IL-10 and TGF-β in the supernatant decreased. MM cells were sensitive to bortezomib (BTZ) induced apoptosis, but SP cells were less sensitive. Co-culture with MM-MSCs significantly reduced BTZ-mediated apoptosis. The addition of α-GA partially restored the sensitivity of MM cells to bortezomib. Conclusion MM-MSCs and multiple myeloma SP cells up-regulate the expression of Cx43 protein, form more GJIC, and promote the proliferation and drug resistance of SP cells by changing the cytokine secretion profile of MSCs, which may be one of the reasons for the recurrence of MM. -
Key words:
- multiple myeloma /
- gap junction connexin /
- tumor microenvironment
-
图 4 MM-MSCs对SP细胞周期的作用
A. RPMI8226单独培养;B. SP细胞单独培养;C. SP+MM-MSCs培养;D. SP+MM-MSCs+α-GA培养
图 5 ND-NSCs及MM-MSCs对SP细胞的体外集落形成的影响
A. SP细胞+ND-MSCs;B. SP细胞+MM-MSCs;C. SP细胞+ND-MSCs+25mmol/L α-GA;D. SP细胞+MM-MSCs+25mmol/L α-GA
图 6 培养上清中细胞因子变化
***P<0.001,与8226组IL-6比较;###P<0.001,与IL-10比较; ▼▼▼P<0.001,与TGF-β比较*P<0.05,**P<0.01,***P<0.001
图 7 RT-PCR分析SP细胞干细胞相关基因表达
A. C-myc;B. KIF4;C. SOX-2;D. Oct-4***P<0.001,与8226组比较;###P<0.001,与SP组比较,*P<0.05,**P<0.01,***P<0.001
图 8 GJ在硼替佐咪诱导的MM细胞凋亡中的作用
A. RPMI8226;B. RPMI8226+BTZ;C. RPMI8226+BTZ+BTZ;D. RPMI8226+BTZ+BTZ+α-GA; E. SP;F. SP+BTZ; G.SP+MM-MSCs+BTZ;H. SP+MM-MSCs+BTZ+α-GA
表 1 干细胞样基因引物序列
基因 引物序列 c-myc 5′CTTCTCTCCGTCCTCGGATTCT
3′GAAGGTGATCCAGACTCTGACCTTKlf-4 5′GCAAGTCCCCTCTCTCCATTA
3′GTAAGGTTTCTCGCCTGTGTGOct-4 5′GGAGATATGCAAAGCAGAAACC
3′CTCAAAATCCTCTCGTTGTGCSox-2 5′CGGCAACCAGAAAAACAGC
3′TCTCCGTCTCCGACAAAAGT 下载: 导出CSV
下载: 导出CSV
表 3 不同培养体系加入阻断剂前后克隆形成能力
组别 单克隆直径
(cm)克隆形成数 克隆形成率
(/2000)SP 0.28±0.16 1722±127 86%±6% SP+ND-MSCs 0.33±0.14 1858±89 93%±4% SP+MM-MSCs 0.38±0.21 1900±85 95%±4% SP+ND-MSCs+GA 0.25±0.22 1532±112 77%±6% SP+MM-MSCs+GA 0.31±0.17 1755±76 88%±4%
下载: 导出CSV
-
[1] BONACQUISTI E E, NGUYEN J. Connexin 43 (Cx43) in cancer: Implications for therapeutic approaches via gap junctions[J]. Cancer Lett,2019,442:439-444. doi: 10.1016/j.canlet.2018.10.043 [2] ZHANG X H, SUN Y, WANG Z Y, et al. Up-regulation of connexin-43 expression in bone marrow mesenchymal stem cells plays a crucial role in adhesion and migration of multiple myeloma cells[J]. Leuk Lymphoma,2015,56(1):211-218. doi: 10.3109/10428194.2014.913289 [3] ZHANG Y M, WANG Z Y, ZHANG L Y, et al. Impact of connexin 43 coupling on survival and migration of multiple myeloma cells[J]. Arch Med Sci,2017,13(6):1335-1346. [4] JAKUBIKOVA J, CHOLUJOVA D, HIDESHIMA T, et al. A novel 3D mesenchymal stem cell model of the multiple myeloma bone marrow niche: biologic and clinical applications[J]. Oncotarget,2016,7(47):77326-77341. doi: 10.18632/oncotarget.12643 [5] GAO M J, KONG Y Y, YANG G, et al. Multiple myeloma cancer stem cells[J]. Oncotarget,2016,7(23):35466-35477. doi: 10.18632/oncotarget.8154 [6] BEHRENS J, KAMERITSCH P, WALLNER S, et al. The carboxyl tail of Cx43 augments p38 mediated cell migration in a gap junction-independent manner[J]. Eur J Cell Biol,2010,89(11):828-838. doi: 10.1016/j.ejcb.2010.06.003 [7] BELLONI D, HELTAI S, PONZONI M, et al. Modeling multiple myeloma-bone marrow interactions and response to drugs in a 3D surrogate microenvironment[J]. Haematologica,2018,103(4):707-716. doi: 10.3324/haematol.2017.167486 [8] XU S, DE VEIRMAN K, DE BECKER A, et al. Mesenchymal stem cells in multiple myeloma: a therapeutical tool or target? Leukemia,2018,32(7):1500-1514. doi: 10.1038/s41375-018-0061-9 [9] ISSA M E, CRETTON S, CUENDET M. Targeting multiple myeloma cancer stem cells with natural products - lessons from other hematological malignancies[J]. Planta Med,2017,83(9):752-760. doi: 10.1055/s-0043-109558 [10] WANG Z Y, YUAN Y Q, ZHANG L Y, et al. Impact of cell fusion in myeloma marrow microenvironment on tumor progression[J]. Oncotarget,2018,9(57):30997-31006. doi: 10.18632/oncotarget.25742 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 8689
- HTML全文浏览量: 3423
- PDF下载量: 15
- 被引次数: 0
