

 下载:
下载:

-
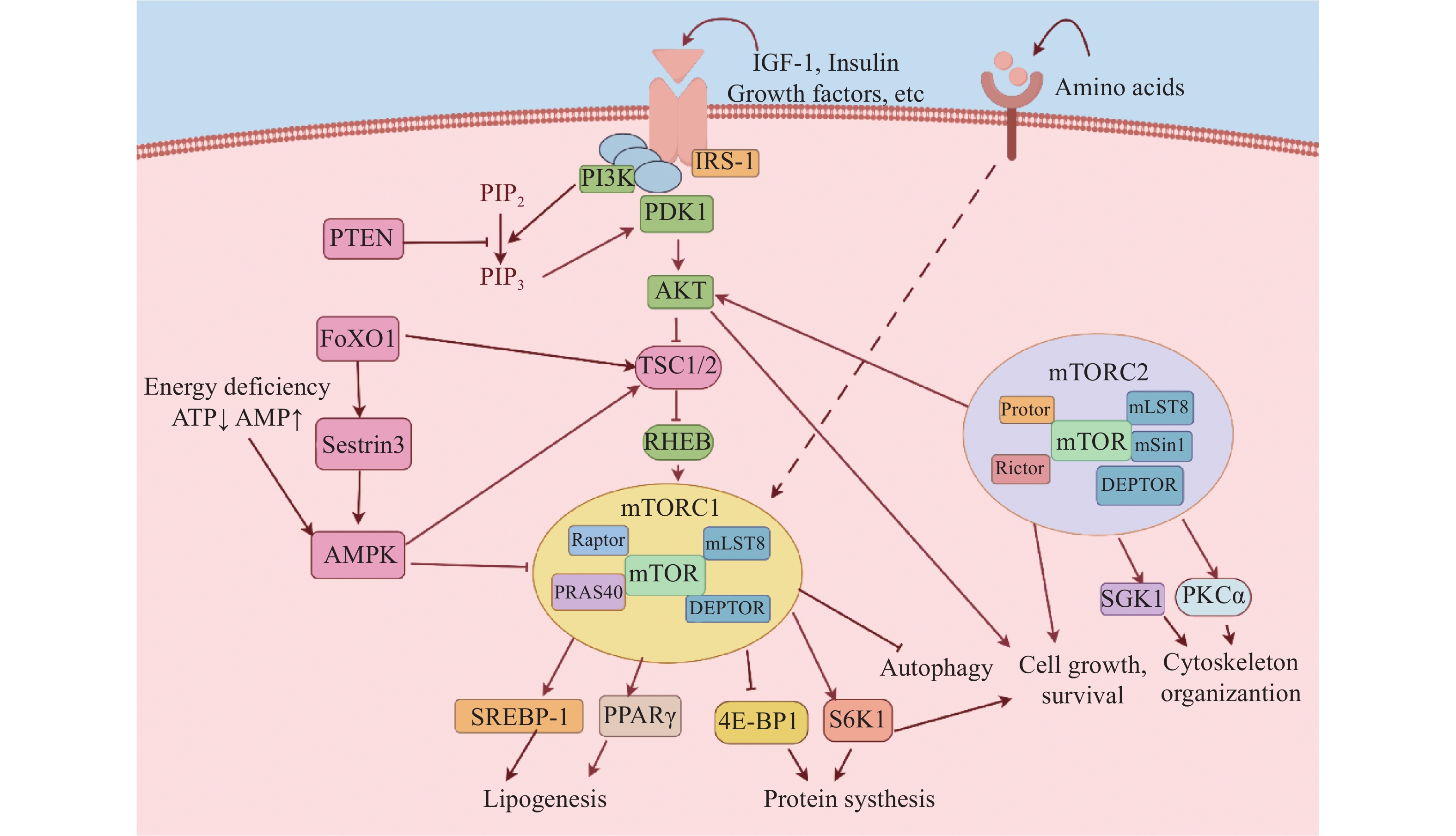
哺乳动物mTOR是磷脂酰肌醇3-激酶(PI3K)家族的丝氨酸/苏氨酸蛋白激酶,是细胞内关键的信号转导枢纽,调节细胞增殖、分化、自噬和代谢等多种生理过程[1]。mTOR通过与不同的蛋白结合形成两种功能不同的复合物:雷帕霉素靶蛋白复合物1(mTORC1)和雷帕霉素靶蛋白复合物2(mTORC2)(图1)。mTORC1由关联调节蛋白(Raptor)、雷帕霉素靶蛋白复合物亚基LST8(mLST8)、40kDa富含脯氨酸的AKT底物(PRAS40)、含DEP结构域的mTOR相互作用蛋白(DEPTOR)组成。mTORC1主要通过感知生长因子、营养物质和细胞能量信号调控下游信号。在生长因子,如胰岛素(Insulin)、胰岛素样生长因子(IGF-1)等激活下,PI3K磷酸化脂酰肌醇-4,5(PIP2)生成PIP3。随后,PIP3招募并结合脂酰肌醇依赖性激酶(PDK1),使蛋白激酶B(AKT)的Thr-308位点磷酸化。激活的AKT磷酸化结节性硬化症复合物1/2(TSC1/2),解除其对脑中富集的Ras同源物(RHEB)的抑制作用,最终激活mTORC1。磷酸酯酶与张力蛋白同源物(PTEN)能抑制PI3K的功能。细胞的能量代谢状态主要通过腺苷单磷酸激活的蛋白激酶(AMPK)调控mTORC1,当细胞能量消耗时,ATP水平降低,导致AMPK被激活,增强TSC2活性而抑制mTORC1。激活的mTORC1磷酸化其下游的核糖体蛋白S6激酶1(S6K1)和真核起始因子4E结合蛋白(4E-BPs)调节mRNA的翻译,从而促进蛋白质合成。此外,mTORC1还通过上调固醇调节元件结合蛋白1(SREBP-1)和过氧化物酶体增殖物激活受体γ(PPARγ)诱导一系列脂肪合成酶表达,进而促进脂质生成。与mTORC1相比,mTORC2的分子调控机制尚未完全明确。目前已知的是,在生长因子刺激下,mTORC2主要受PI3K-AKT信号通路调控。Insulin诱导的PI3K可通过促进mTORC2与核糖体相互作用来激活mTORC2。当mTORC2被激活时,它会磷酸化AKT的Ser473位点,这是AKT完全激活所必需的。这些分子事件最终参与调节细胞生长、存活和肌动蛋白骨架等过程(图1)。
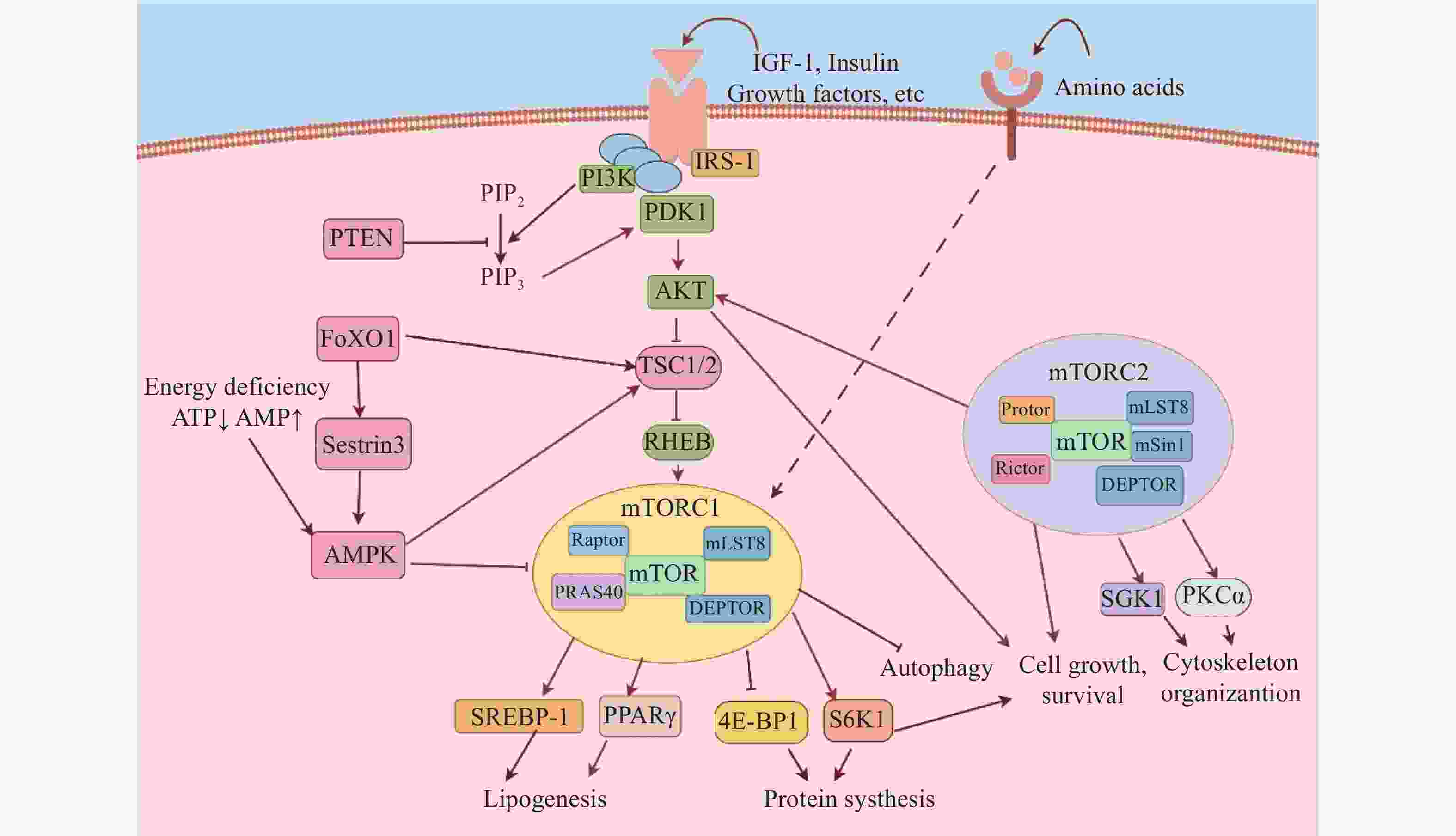
图 1 mTOR信号通路转导
-
mTOR信号在皮肤形态发生和稳态调控中起着关键作用,其激活可增强细胞合成代谢并抑制分解代谢,进而调控蛋白质合成和脂质代谢相关基因的表达。痤疮患者皮肤中mTOR的表达水平显著高于健康皮肤,同时,在痤疮皮损发生过程中,mTOR和S6K1的磷酸化水平持续增加,提示mTOR信号通路的过度活化在痤疮病理进程中可能起着重要作用[2]。本文我们重点综述了mTOR信号通路调控痤疮发生发展的潜在机制,并总结了靶向调控mTOR信号在治疗痤疮的最新进展。
-
寻常痤疮(AV)是一种发生于毛囊皮脂腺单位的慢性炎症性皮肤病,涉及皮脂腺过度分泌皮脂、皮脂腺导管上皮过度角化、痤疮丙酸杆菌(C.acne)异常定植,以及炎症介质的产生和释放等病理过程[3]。AV的发生与遗传、环境因素(如饮食和气候)、激素(如雄激素和IGF-1)以及微生态失衡等因素有关。
IGF-1在痤疮的发病机制中起关键作用[4]。研究显示,血清IGF-1含量与痤疮皮损数量和皮脂分泌呈正相关[4]。同时,IGF-1在痤疮患者的表皮和毛囊皮脂腺单位中表达显著增加。有趣的是,先天性缺乏IGF-1的侏儒症(Laron综合征)患者几乎不患痤疮,但以高剂量IGF-1治疗会诱发痤疮,而降低IGF-1的剂量或停用IGF-1则使痤疮消退[5]。粉刺是痤疮的早期表现形式。研究显示,IGF-1可通过IGF-1受体(IGF-1R)激活PI3K/AKT/mTORC1信号通路,进而诱导毛囊漏斗部和皮脂腺导管角质形成细胞增殖,并促进皮脂腺细胞的增殖、分化和脂质生成,最终诱导粉刺形成[6]。相应地,抑制IGF-1信号可缓解痤疮症状,如脂质生长、角质分化异常等[7]。
Insulin作用机制与IGF-1相似,通过作用于胰岛素受体(IRS-1)和IGF-1R激活PI3K/AKT/mTOR通路诱发痤疮[4]。低分化的皮脂腺细胞对Insulin刺激更加敏感[9]。胰岛素抵抗(IR)是AV发展的独立致病因素[10]。IR在痤疮患者中更为常见,而mTORC1信号转导增加则可能是IR和痤疮发生之间的关键节点。mTORC1激活的S6K1可磷酸化IRS-1,抑制Insulin信号转导,引起IR和高胰岛素血症[9]。高水平的胰岛素继而激活皮脂腺细胞的mTOR信号通路,增加皮脂合成,最终诱发痤疮。多囊卵巢综合征、高雄激素血症、黑棘皮病患者也常伴有高胰岛素血症,并表现为皮脂分泌过多和痤疮[8,10]。
SREBP-1是调节脂质生成的关键转录因子。研究显示,mTORC1可通过调控SREBP-1表达促进脂合成。mTORC1可通过S6K1激活SREBP切割激活蛋白(SCAP),进而使SREBP-1以活性形式进入核内,上调脂质合成相关基因表达。Lipin-1是SREBP的负调控因子[11]。mTORC1还可磷酸化并抑制Lipin-1激活,解除Lipin-1对SREBP-1的抑制[11]。在营养过剩或高Insulin/IGF-1条件下,SREBP-1被激活;而低血糖负荷饮食后,痤疮患者皮肤中SREBP-1表达显著降低。C.acne 异常是导致痤疮发展的关键因素。研究显示,在C.acne和IGF-1/Insulin诱导的痤疮细胞模型中,SREBP-1的表达显著增加,而抑制SREBP-1的表达能显著抑制皮脂生成[7,12]。除脂质合成外,SREBP-1还可通过调节皮脂中单不饱和脂肪酸比例[13],促进炎症应答,如SREBP-1调节的油酸可诱导角质形成细胞异常分化并释放IL-1α,引起皮脂腺炎症,进而诱发痤疮[4]。
叉头框蛋白O1(FoxO1)是另一个调控mTORC1信号的关键转录因子,能通过诱导应激响应蛋白Sestrin3表达激活AMPK,从而抑制mTORC1[14]。FoxO1与痤疮发生发展有密切关系[4]。IGF-1/Insulin激活PI3K/AKT通路使FoxO1磷酸化并转移至胞质,解除TSC1/TSC2的抑制,激活mTORC1促进脂质合成[15]。FoxO1还可抑制SREBP-1c、PPARγ和肝X受体α(LXRα)等一系列脂质生成转录因子活性,进而调控细胞内脂质稳态。IGF-1还可通过PI3K/AKT/FoxO1通路上调皮脂腺细胞中Toll样受体2/4(TLR2/4)的表达水平,促进IL-1、IL-6、IL-8等促炎细胞因子的产生,加剧痤疮的炎症反应[16]。在痤疮患者皮肤中,FoxO1表达水平降低,同时其向胞质转移增多,并且和血清IGF-1水平增高相关[3,16]。这些研究提示,IGF-1/FoxO1或许在痤疮发生发展中发挥作用,这有待进一步探索。综上,Insulin/IGF-1不仅通过激活mTORC1通路增加皮脂从头合成,还可诱导炎症反应,促进痤疮进展。
-
大量流行病学调查显示高碳水化合物饮食(高糖饮食)和高乳制品(奶制品、巧克力等)饮食与痤疮呈正相关。高糖饮食和奶制品升高餐后Insulin/IGF-1水平,激活mTORC1信号,进而诱发或加剧痤疮[14,17]。相反,低糖饮食降低IGF-1水平,缓解痤疮症状,并且高乳饮食还可给机体提供大量氨基酸,如亮氨酸和谷氨酰胺,进而调控mTORC1的活性[18]。比如,亮氨酸是Insulin激活mTORC1活性所必需的氨基酸[19],谷氨酰胺与亮氨酸协同激活mTORC1[20]。皮脂腺细胞缺乏谷氨酰胺会导致增殖减慢和皮脂生成减少,提示谷氨酰胺在皮脂生成中具有重要作用[21],这或许与mTOC1活性相关。新近研究显示,牛乳和母乳中含有丰富外泌体,这些外泌体富含的生物活性分子可调节mTOR通路活性。例如,牛奶外泌体中通常含有的microRNA-21可抑制多种mTOR通路负调节因子的基因表达,包括PTEN和FoxO1等,激活mTOR信号,最终加剧皮脂分泌,促进痤疮发生[6]。
-
雄激素在痤疮发生发展中有重要作用[22]。雄激素水平与痤疮发病时期(青春期)高度吻合。此外,患有与雄激素过多相关疾病的患者患痤疮的概率更高,如多囊卵巢综合征和先天性肾上腺皮质增生症,而雄激素缺乏或不足患者往往不会患痤疮。雄激素受体(AR)在皮脂腺中广泛表达,而痤疮易发区域AR表达较高,以上均表明雄激素在痤疮进展中的重要作用。
雄激素可通过多环节影响痤疮进展。研究显示,雄激素睾酮和二氢睾酮(DHT)促进皮脂腺细胞增殖及脂合成相关酶表达。DHT还能上调皮脂腺细胞中TNF-α、IL-6等促炎细胞因子的表达,并通过促进成纤维细胞生长因子10(FGF10)分泌影响角质形成细胞的增殖分化[23]。
雄激素和mTOR信号通路存在多个层面的交叉调控。雄激素可提高男性系统或局部IGF-1水平,进而间接激活皮肤细胞内的PI3K/AKT/mTOR通路。反之,IGF-1/Insulin可诱导雄激素合成,并且通过增强5α-还原酶活性促进睾酮转化为活性更强的DHT[8]。更重要的是,IGF-1//Insulin可通过抑制PI3K/AKT/FoxO1信号轴来激活AR信号[8,11]。因此,IGF-1/mTORC1与雄激素的正向反馈调节是痤疮进展的重要因素。
-
目前,临床上治疗痤疮最有效的口服药物是异维甲酸,外用药主要是阿达帕林凝胶和过氧苯甲酰凝胶,其他常用的药物还有壬二酸、大环内酯类和四环素类抗生素等。研究表明多种痤疮治疗药物可能通过激活FoxO1和抑制mTORC1发挥治疗作用,如异维甲酸可诱导FoxO1基因表达,并提高痤疮患者皮脂腺细胞核内FoxO1水平;过氧化苯甲酰通过提高胞内ROS水平上调核FoxO1水平,进而通过诱导Sestrin3表达激活负调控mTORC1的AMPK;壬二酸通过抑制线粒体呼吸链增加ROS生成,进而上调FoxOs和Sestrin3表达以及胞内AMP水平,从而激活AMPK;四环素、红霉素和其他大环内酯类抗生素可通过抑制NF-κB信号通路发挥抗炎作用,同时也可能会增强TSC1/TSC2复合物对mTORC1的抑制活性[11]。近年的研究发现治疗2型糖尿病的经典药物二甲双胍对痤疮也有治疗作用,其药理机制可能通过激活AMPK抑制mTORC1,并且能降低血清IGF-1水平,从而减少皮脂分泌和减轻炎症[24]。这些研究表明痤疮治疗药物可通过不同的机制抑制mTOR通路,从而减少皮脂分泌、抑制皮脂腺导管异常角化和减弱炎症反应,同时也提示mTOR信号通路有可能成为痤疮治疗的新靶标。
近年来越来越多的天然活性成分被发现通过抑制mTOR信号通路发挥痤疮治疗作用。没食子儿茶素没食子酸酯(EGCG)被发现是PI3K/mTOR的双重抑制剂[25]。EGCG可通过阻断AKT/mTOR信号转导抑制IGF-1刺激的SZ95人皮脂腺细胞生成脂质,并抑制IL-1、IL-6和IL-8等炎症因子表达[16]。EGCG还可通过激活AMPK抑制mTORC1/SREBP-1信号通路以减少皮脂生成。EGCG外用可明显减小兔耳皮脂腺的体积和粉刺大小[16],并且在一项为期8周的随机分组临床试验中显著改善痤疮[26]。
白藜芦醇也能下调PI3K/AKT/mTORC1信号。它可通过阻断PI3K/AKT信号通路抑制SZ95人皮脂腺细胞生长,并且通过激活AMPK信号通路抑制亚油酸诱导的SZ95人皮脂腺细胞的脂肪生成和炎症应答[27]。除抑制脂质生成和抗炎作用外,白藜芦醇还可抑制痤疮丙酸杆菌的生长,阻止其生物膜的形成,而且白藜芦醇凝胶局部治疗可显著减少痤疮患者面部的微粉刺、丘疹和脓疱的数量。
大黄素是一种存在于大黄和虎杖等药用植物中的蒽醌类化合物。大黄素能通过抑制PI3K/AKT/FoxO1通路抑制IGF-1诱导的皮脂腺脂质生成,并且与异维甲酸有协同作用[28]。此外,大黄素还能够减少痤疮丙酸杆菌刺激的皮脂腺细胞和角质形成细胞中IL-1β、IL-6和IL-8等促炎细胞因子的表达,抑制角质形成细胞的过度增值,因而对痤疮可能具有治疗作用[28]。
除了上述天然化合物之外,还有多种其他的天然成分也被发现具有调节皮脂腺细胞的mTOR信号通路和皮脂生成的作用,而且这些天然成分大多也同时具有抗炎或抑菌活性,有潜在的痤疮治疗作用(表1)。
表 1 具有调节皮脂腺细胞mTOR信号通路作用的其他天然成分
中文名 英文名 来源 主要药理作用 参考文献 蜂毒素 Melittin 蜂毒 抑制IGF-1R/AKT/mTOR/SREBP-1信号通路,抑制IGF-1诱导的人皮脂腺细胞的脂质生成和促炎细胞因子的表达;在小鼠痤疮模型上通过阻断PI3K/mTOR信号通路抑制抑制痤疮丙酸杆菌诱导的皮脂生成和炎症反应,缓解痤疮皮损症状。 [7] 绿原酸 Chlorogenic acid 金银花、咖啡豆等 下调皮脂腺细胞中AKT/mTOR/FoxO1/SREBP-1信号通路,抑制脂质生成;缓解痤疮丙酸杆菌诱导的小鼠耳部肿胀和发红等痤疮症状,抑制痤疮丙酸杆菌刺激的皮脂腺细胞和HaCaT细胞中促炎细胞因子表达。 [29] 大麻籽己烷提取物 Hemp seed hexane extracts 大麻籽 调节AMPK和AKT/FoxO1信号通路,抑制IGF-1诱导的人皮脂腺细胞的脂质生成和炎症应答;抑制痤疮丙酸杆菌的增殖及其在HaCaT细胞中引起的炎症应答。 [30] 欧前胡素 Imperatorin 白芷 阻断AKT/SREBP-1通路,抑制IGF-1刺激的皮脂腺细胞合成皮脂。 [31] 银杏叶素 Bilobetin 银杏叶 阻断AKT/SREBP-1通路,抑制IGF-1刺激的皮脂腺细胞合成皮脂。 [32] 桔梗皂苷D Platycodin D 桔梗 阻断皮脂腺细胞内的IGF-1R/PI3K/AKT/SREBP-1通路,抑制皮脂生成;抑制LPS刺激的角质形成细胞分泌促炎细胞因子。 [33] 羽扇豆醇 Lupeol 龙葵 阻断IGF-1R/PI3K/AKT/SREBP-1通路,抑制皮脂腺细胞的脂质生成,抑制痤疮丙酸杆菌刺激的皮脂腺细胞和HaCaT细胞的炎症应答;降低粉刺或皮脂腺周围浸润炎症细胞数量和IGF-1R、SREBP-1、TLR2、NF-κB p65、IL-8、IL-1α等蛋白水平。 [34] -
mTOR信号通路是调控皮脂腺脂质生成、皮脂细胞增殖分化和炎症应答的关键通路,其功能失调在痤疮发生发展中起重要作用,因而已成为痤疮治疗的新靶标。尽管新近的研究让我们对mTOR信号轴和痤疮的关系有了新的认识,但这仅仅是一个开始,mTOR信号轴到底是如何影响痤疮发展,其下游关键的信号分子如何,仍需进一步探索。此外,迄今已发现多种天然成分可通过靶向mTOR通路调节皮脂腺细胞脂质生成并抑制炎症反应,有望成为辅助或替代药物用于痤疮防治,但大多仍处于候选药物研发阶段,尚需开展进一步的临床前研究和临床试验评价其疗效和安全性。鉴于mTOR信号通路在痤疮病理进程中的重要作用,进一步深入探讨如何精准靶向调控这一关键通路将为痤疮防治提供新的靶点和治疗选择。
Research progress on acne pathogenesis and medicine intervention based on mTOR signaling pathway
-
摘要: 雷帕霉素靶蛋白(mTOR)是细胞内重要的信号转导枢纽,调节细胞生长、增殖和代谢等过程。mTOR信号通路的功能失调与多种炎症和过度增殖性皮肤病有关,尤其在痤疮的病理机制中扮演重要角色。本文综述了mTOR信号通路在痤疮中的调控机制以及作为治疗靶标的研究进展。Abstract: The mechanistic target of rapamycin (mTOR) is an important signaling functions as a central regulator of cell growth, proliferation and metabolism. Dysregulated mTOR signaling has been implicated in various inflammatory and hyperproliferative skin conditions. Emerging data suggest that mTOR signaling is tightly associated with acne pathogenesis. The regulatory mechanisms of the mTOR signaling pathway in the pathogenesis of acne were summarized and the current progress of therapeutic targeting mTOR pathway for acne were discussed in this paper.
-
Key words:
- mTOR /
- Acne /
- Sebaceous glands /
- Lipogenesis /
- Therapy
-
表 1 具有调节皮脂腺细胞mTOR信号通路作用的其他天然成分
中文名 英文名 来源 主要药理作用 参考文献 蜂毒素 Melittin 蜂毒 抑制IGF-1R/AKT/mTOR/SREBP-1信号通路,抑制IGF-1诱导的人皮脂腺细胞的脂质生成和促炎细胞因子的表达;在小鼠痤疮模型上通过阻断PI3K/mTOR信号通路抑制抑制痤疮丙酸杆菌诱导的皮脂生成和炎症反应,缓解痤疮皮损症状。 [7] 绿原酸 Chlorogenic acid 金银花、咖啡豆等 下调皮脂腺细胞中AKT/mTOR/FoxO1/SREBP-1信号通路,抑制脂质生成;缓解痤疮丙酸杆菌诱导的小鼠耳部肿胀和发红等痤疮症状,抑制痤疮丙酸杆菌刺激的皮脂腺细胞和HaCaT细胞中促炎细胞因子表达。 [29] 大麻籽己烷提取物 Hemp seed hexane extracts 大麻籽 调节AMPK和AKT/FoxO1信号通路,抑制IGF-1诱导的人皮脂腺细胞的脂质生成和炎症应答;抑制痤疮丙酸杆菌的增殖及其在HaCaT细胞中引起的炎症应答。 [30] 欧前胡素 Imperatorin 白芷 阻断AKT/SREBP-1通路,抑制IGF-1刺激的皮脂腺细胞合成皮脂。 [31] 银杏叶素 Bilobetin 银杏叶 阻断AKT/SREBP-1通路,抑制IGF-1刺激的皮脂腺细胞合成皮脂。 [32] 桔梗皂苷D Platycodin D 桔梗 阻断皮脂腺细胞内的IGF-1R/PI3K/AKT/SREBP-1通路,抑制皮脂生成;抑制LPS刺激的角质形成细胞分泌促炎细胞因子。 [33] 羽扇豆醇 Lupeol 龙葵 阻断IGF-1R/PI3K/AKT/SREBP-1通路,抑制皮脂腺细胞的脂质生成,抑制痤疮丙酸杆菌刺激的皮脂腺细胞和HaCaT细胞的炎症应答;降低粉刺或皮脂腺周围浸润炎症细胞数量和IGF-1R、SREBP-1、TLR2、NF-κB p65、IL-8、IL-1α等蛋白水平。 [34]  下载: 导出CSV
下载: 导出CSV
-
[1] LIU X J, GUO B, LI Q Y, et al. mTOR in metabolic homeostasis and disease[J]. Exp Cell Res, 2024, 441(2): 114173. doi: 10.1016/j.yexcr.2024.114173 [2] AKDAĞ N, ATLI E, ZHURI D, et al. A Study of FoxO1, mTOR, miR-21, miR-29b, and miR-98 Expression Levels Regarding Metabolic Syndrome in Acne Vulgaris Patients[J]. Cureus, 2024, 16(3): e56562. doi: 10.7759/cureus.56562 [3] KIM H J, KIM Y H. Exploring acne treatments: from pathophysiological mechanisms to emerging therapies[J]. Int J Mol Sci, 2024, 25(10): 5302. doi: 10.3390/ijms25105302 [4] MELNIK B C. Acne transcriptomics: fundamentals of acne pathogenesis and isotretinoin treatment[J]. Cells, 2023, 12(22): 2600. doi: 10.3390/cells12222600 [5] BEN-AMITAI D, LARON Z. Effect of insulin-like growth factor-1 deficiency or administration on the occurrence of acne[J]. J Eur Acad Dermatol Venereol, 2011, 25(8): 950-954. doi: 10.1111/j.1468-3083.2010.03896.x [6] MELNIK B C. Linking diet to acne metabolomics, inflammation, and comedogenesis: an update[J]. Clin Cosmet Investig Dermatol, 2015, 8: 371-388. doi: 10.2147/ccid.s69135 [7] GU H, AN H J, GWON M G, et al. Bee venom and its major component melittin attenuated Cutibacterium acnes- and IGF-1-induced acne vulgaris via inactivation of Akt/mTOR/SREBP signaling pathway[J]. Int J Mol Sci, 2022, 23(6): 3152. doi: 10.3390/ijms23063152 [8] OKORO O E, CAMERA E, FLORI E, et al. Insulin and the sebaceous gland function[J]. Front Physiol, 2023, 14: 1252972. doi: 10.3389/fphys.2023.1252972 [9] OTTAVIANI M, FLORI E, MASTROFRANCESCO A, et al. Sebocyte differentiation as a new target for acne therapy: an in vivo experience[J]. J Eur Acad Dermatol Venereol, 2020, 34(8): 1803-1814. doi: 10.1111/jdv.16252 [10] GRUSZCZYŃSKA M, SADOWSKA-PRZYTOCKA A, SZYBIAK W, et al. Insulin resistance in patients with acne vulgaris[J]. Biomedicines, 2023, 11(8): 2294. doi: 10.3390/biomedicines11082294 [11] PANWAR V, SINGH A, BHATT M, et al. Multifaceted role of mTOR(mammalian target of rapamycin)signaling pathway in human health and disease[J]. Signal Transduct Target Ther, 2023, 8(1): 375. [12] GU H, AN H J, GWON M G, et al. The effects of synthetic SREBP-1 and PPAR-γ decoy oligodeoxynucleotide on acne-like disease in vivo and in vitro via lipogenic regulation[J]. Biomolecules, 2022, 12(12): 1858. doi: 10.3390/biom12121858 [13] CAO K, LIU Y, LIANG N N, et al. Fatty acid profiling in facial sebum and erythrocytes from adult patients with moderate acne[J]. Front Physiol, 2022, 13: 921866. doi: 10.3389/fphys.2022.921866 [14] MOON K M, LEE M K, HWANG T, et al. The multi-functional roles of forkhead box protein O in skin aging and diseases[J]. Redox Biol, 2021, 46: 102101. doi: 10.1016/j.redox.2021.102101 [15] RYGUŁA I, PIKIEWICZ W, KAMINIÓW K. Impact of diet and nutrition in patients with acne vulgaris[J]. Nutrients, 2024, 16(10): 1476. doi: 10.3390/nu16101476 [16] IM M, KIM S Y, SOHN K C, et al. Epigallocatechin-3-gallate suppresses IGF-I-induced lipogenesis and cytokine expression in SZ95 sebocytes[J]. J Invest Dermatol, 2012, 132(12): 2700-2708. doi: 10.1038/jid.2012.202 [17] BALDWIN H, TAN J. Effects of diet on acne and its response to treatment[J]. Am J Clin Dermatol, 2021, 22(1): 55-65. doi: 10.1007/s40257-020-00542-y [18] GOUL C, PERUZZO R, ZONCU R. The molecular basis of nutrient sensing and signalling by mTORC1 in metabolism regulation and disease[J]. Nat Rev Mol Cell Biol, 2023, 24(12): 857-875. doi: 10.1038/s41580-023-00641-8 [19] POLIDORI N, GRASSO E A, CHIARELLI F, et al. Amino acid-related metabolic signature in obese children and adolescents[J]. Nutrients, 2022, 14(7): 1454. doi: 10.3390/nu14071454 [20] YOSHIMURA R, NOMURA S. Co-ingestion of glutamine and leucine synergistically promotes mTORC1 activation[J]. Sci Rep, 2022, 12(1): 15870. doi: 10.1038/s41598-022-20251-2 [21] DOWNIE M T, KEALEY T. Human sebaceous glands engage in aerobic glycolysis and glutaminolysis[J]. Br J Dermatol, 2004, 151(2): 320-327. doi: 10.1111/j.1365-2133.2004.06004.x [22] HU T T, WEI Z Y, JU Q, et al. Sex hormones and acne: state of the art[J]. J Dtsch Dermatol Ges, 2021, 19(4): 509-515. doi: 10.1111/ddg.14426 [23] GRATTON R, DEL VECCHIO C, ZUPIN L, et al. Unraveling the role of sex hormones on keratinocyte functions in human inflammatory skin diseases[J]. Int J Mol Sci, 2022, 23(6): 3132. doi: 10.3390/ijms23063132 [24] SZEFLER L, SZYBIAK-SKORA W, SADOWSKA-PRZYTOCKA A, et al. Metformin therapy for acne vulgaris: a meta-analysis[J]. Pharmaceuticals, 2024, 17(6): 728. [25] VAN ALLER G S, CARSON J D, TANG W, et al. Epigallocatechin gallate(EGCG), a major component of green tea, is a dual phosphoinositide-3-kinase/mTOR inhibitor[J]. Biochem Biophys Res Commun, 2011, 406(2): 194-199. [26] YOON J Y, KWON H H, MIN S U, et al. Epigallocatechin-3-gallate improves acne in humans by modulating intracellular molecular targets and inhibiting P. acnes[J]. J Invest Dermatol, 2013, 133(2): 429-440. doi: 10.1038/jid.2012.292 [27] WEI Z Y, CHEN G J, HU T T, et al. Resveratrol ameliorates lipid accumulation and inflammation in human SZ95 sebocytes via the AMPK signaling pathways in vitro[J]. J Dermatol Sci, 2021, 103(3): 156-166. doi: 10.1016/j.jdermsci.2021.07.010 [28] LIU S, LUO X H, LIU Y F, et al. Emodin exhibits anti-acne potential by inhibiting cell growth, lipogenesis, and inflammation in human SZ95 sebocytes[J]. Sci Rep, 2023, 13(1): 21576. doi: 10.1038/s41598-023-48709-x [29] LUO J, HE W Y, LI X, et al. Anti-acne vulgaris effects of chlorogenic acid by anti-inflammatory activity and lipogenesis inhibition[J]. Exp Dermatol, 2021, 30(6): 865-871. doi: 10.1111/exd.14277 [30] JIN S, LEE M Y. The ameliorative effect of hemp seed hexane extracts on the Propionibacterium acnes-induced inflammation and lipogenesis in sebocytes[J]. PLoS One, 2018, 13(8): e0202933. doi: 10.1371/journal.pone.0202933 [31] HWANG Y L, IM M, LEE M H, et al. Inhibitory effect of imperatorin on insulin-like growth factor-1-induced sebum production in human sebocytes cultured in vitro[J]. Life Sci, 2016, 144: 49-53. doi: 10.1016/j.lfs.2015.11.027 [32] WANG C, HWANG Y L, LI X M, et al. Inhibition of insulin-like growth factor-1-induced sebum production by bilobetin in cultured human sebocytes[J]. Ann Dermatol, 2019, 31(3): 294-299. doi: 10.5021/ad.2019.31.3.294 [33] SUH Y, YANG J H, YOON J Y, et al. Platycodin D may improve acne and prevent scarring by downregulating SREBP-1 expression via inhibition of IGF-1R/PI3K/Akt pathway and modulating inflammation with an increase in collagen[J]. Ann Dermatol, 2018, 30(5): 581-587. doi: 10.5021/ad.2018.30.5.581 [34] KWON H H, YOON J Y, PARK S Y, et al. Activity-guided purification identifies lupeol, a pentacyclic triterpene, as a therapeutic agent multiple pathogenic factors of acne[J]. J Invest Dermatol, 2015, 135(6): 1491-1500. doi: 10.1038/jid.2015.29 -

 点击查看大图
点击查看大图
图(1) / 表(1)
计量
- 文章访问数: 752
- HTML全文浏览量: 293
- PDF下载量: 2
- 被引次数: 0
