

 下载:
下载:



-
雷公藤为卫矛科雷公藤属植物雷公藤Tripterygium wilfordii Hook.f.的干燥根,是一种公认的同时具有较强药效和较强毒性的中药材,广泛用于治疗类风湿性关节炎、肾病综合征、系统性红斑狼疮等疾病[1]。但其毒性较大,不良反应高发且严重,常使患者不能耐受[2]。雷公藤的主要有效成分为二萜类、三萜类和生物碱类,研究表明各种成分均具有不同程度的抗炎、抗肿瘤和免疫抑制等活性[3-4],其中生物碱的毒性要小于二萜和三萜类成分,临床应用具有疗效好,不良反应较小的特点[5]。雷公藤次碱是雷公藤生物碱中含量较高的一种倍半萜类单体化合物。目前研究显示其具有良好的杀虫活性,其他药理活性和机理研究较少,特别是抗炎和免疫抑制活性方面。
本研究主要通过建立脂多糖(LPS)诱导的RAW264.7细胞炎症模型,探讨雷公藤次碱对LPS诱导的炎性因子NO,IL-1β,TNF-α和IL-6释放的影响,用免疫印迹法考察雷公藤次碱对TRAF6、IRAK、NF-κB、IκBα、JNK、ERK、p38等关键蛋白表达或磷酸化的影响,探讨其可能的抗炎作用机制,为促进雷公藤次碱的应用研究提供基础。
-
雷公藤次碱(纯度98%,上海纯优生物科技有限公司);DMEM细胞培养基、胎牛血清(Life Technologies GIBCO);Griess试剂、细胞计数盒-8 (CCK-8)(上海碧云天生物技术有限公司);脂多糖(LPS, Sigma-Aldrich);IL-1β、TNF-α和IL-6检测试剂盒(达科为生物技术有限公司);核质提取试剂盒(Dojindo Laboratories);TRAF6、IRAF、p-IRAF、p-p38、p38、p-JNK、JNK、p-ERK、ERK、p-IκBα、IκBα、NF-κB p65、β-actin、HRP山羊抗兔抗体(Cell Signaling Technology);RAW264.7细胞购自中国科学院上海细胞库。
-
RAW264.7细胞在含10%的新生小牛血清及100 U/ml青霉素、链霉素的DMEM高糖培养液中进行培养,培养条件为5%CO2、37 ℃,隔天换液,每日观察细胞的生长状况。实验时取对数生长期RAW264.7细胞,胰酶消化,加入新鲜培养基反复吹打成细胞混悬液,细胞增殖实验、细胞因子分泌含量测定调整细胞浓度为1×105 cells/ml,以每孔100 μl细胞悬液接种于96孔细胞培养板中,细胞蛋白表达测定调整细胞浓度到2.25×105 cells/ml,以每孔2 ml接种于6孔细胞培养板中,然后细胞置于37 ℃、5%CO2培养箱中培养过夜。在药物处理前2 h给予LPS溶液(使其终浓度为1 μg/ml)诱导RAW264.7细胞发生炎症反应建立模型。设空白组(不给予LPS和药物处理)、模型组(给予LPS处理)、药物组(给予LPS诱导后再给予相应浓度的药物进行处理),每组设6个复孔(n=6)。
-
设空白组和药物组,药物组分别给予终浓度为200、100、50和25 μmol/L的雷公藤次碱药液进行处理。细胞分别培养24和48 h,实验结束4 h前加入10 μl CCK-8试剂,结束后以酶标仪于450 nm处检测吸光度(A)。
-
设空白组、模型组和药物组,药物组细胞给予LPS诱导后分别给予终浓度为100、50、25 μmol/L的药液进行处理。细胞培养24 h后取出,吸取细胞上清液,Griess试剂法检测上清中NO光密度值,ELISA试剂盒检测上清中IL-1β、TNF-α和IL-6的光密度值。
-
设空白组、模型组和药物组,药物组细胞给予LPS诱导后分别给予终浓度为100、50、25 μmol/L的药液进行处理。细胞培养24 h后取出,用4 ℃预冷的PBS液洗涤各孔两次,然后,吸弃PBS液,每孔中加入200 μl 4 ℃预冷的RIPA提取细胞蛋白,在13000 r/min下离心20 min,弃去上清,得各样本。用Bradford法对蛋白定量;使用细胞核/质提取试剂盒制备用于检测p65/NF-κB的胞质和核提取物。每孔加入20 μg蛋白上样,在SDS-PAGE凝胶中电泳,蛋白转移到PVDF膜上,用BSA封闭2 h。分别用相应一抗4 ℃孵育过夜,然后用与辣根过氧化物酶偶联的二抗(1∶1000)孵育2 h。最后用 ECL超敏发光液显影,于凝胶成像仪中曝光,使用 LAS成像软件对条带进行定量分析
-
实验数据的统计分析用(
$ \bar x \pm s $ )表示,采用单因素方差分析法进行统计学处理。应用Alpha软件处理Western Blot实验所得目标带的光密度值,并以各自的β-actin蛋白条带灰度值与目标条带灰度值的比值作为蛋白相对表达水平值。 -
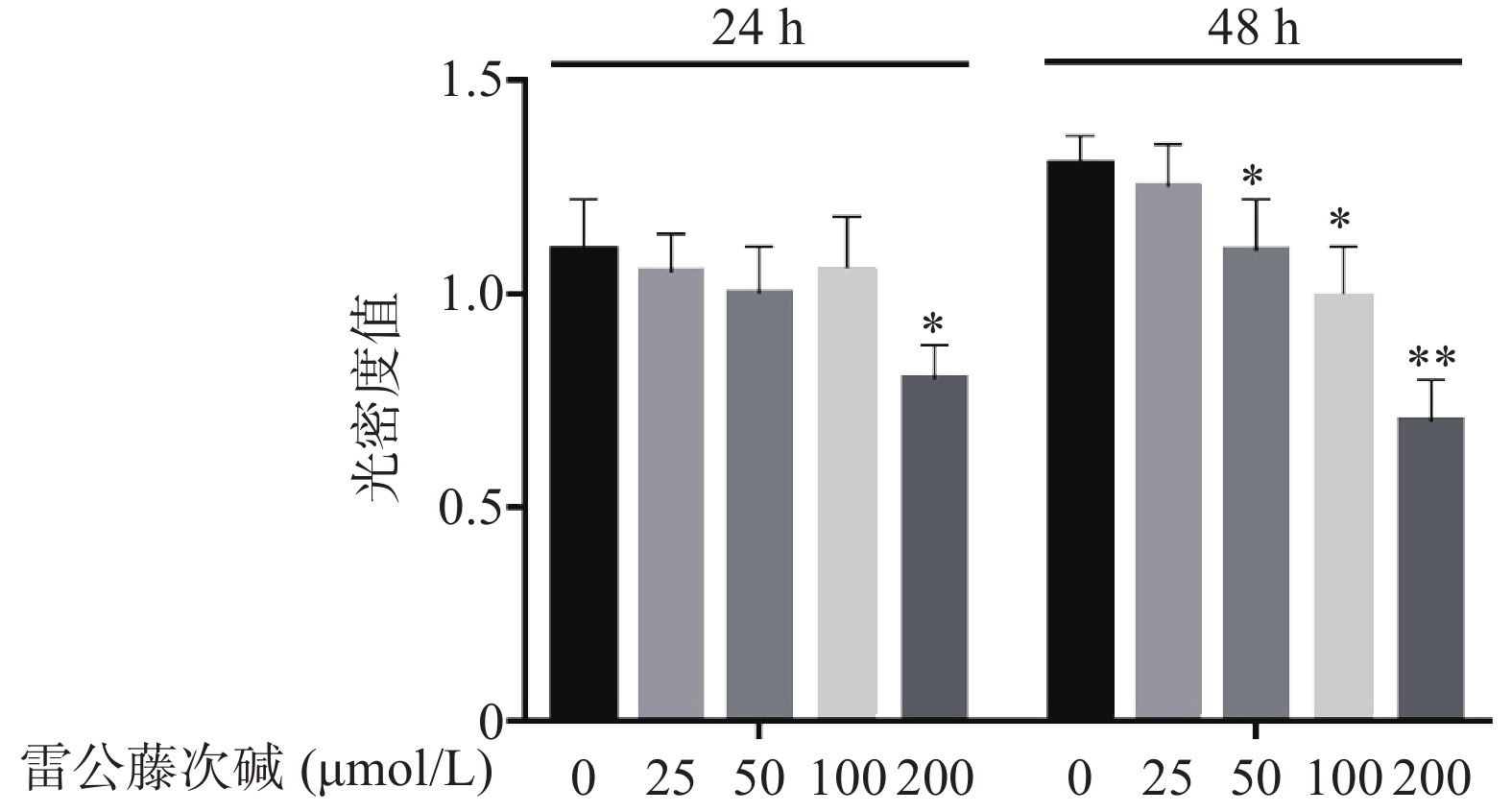
图1显示了不同浓度雷公藤次碱对RAW264.7细胞的细胞毒性作用。给予雷公藤次碱(25~200 μmol/L)孵育24 h后,200 μmol/L雷公藤次碱可显著抑制RAW264.7细胞活性;孵育48 h后,50、100和200 μmol/L雷公藤次碱均可显著抑制RAW264.7细胞的活性。
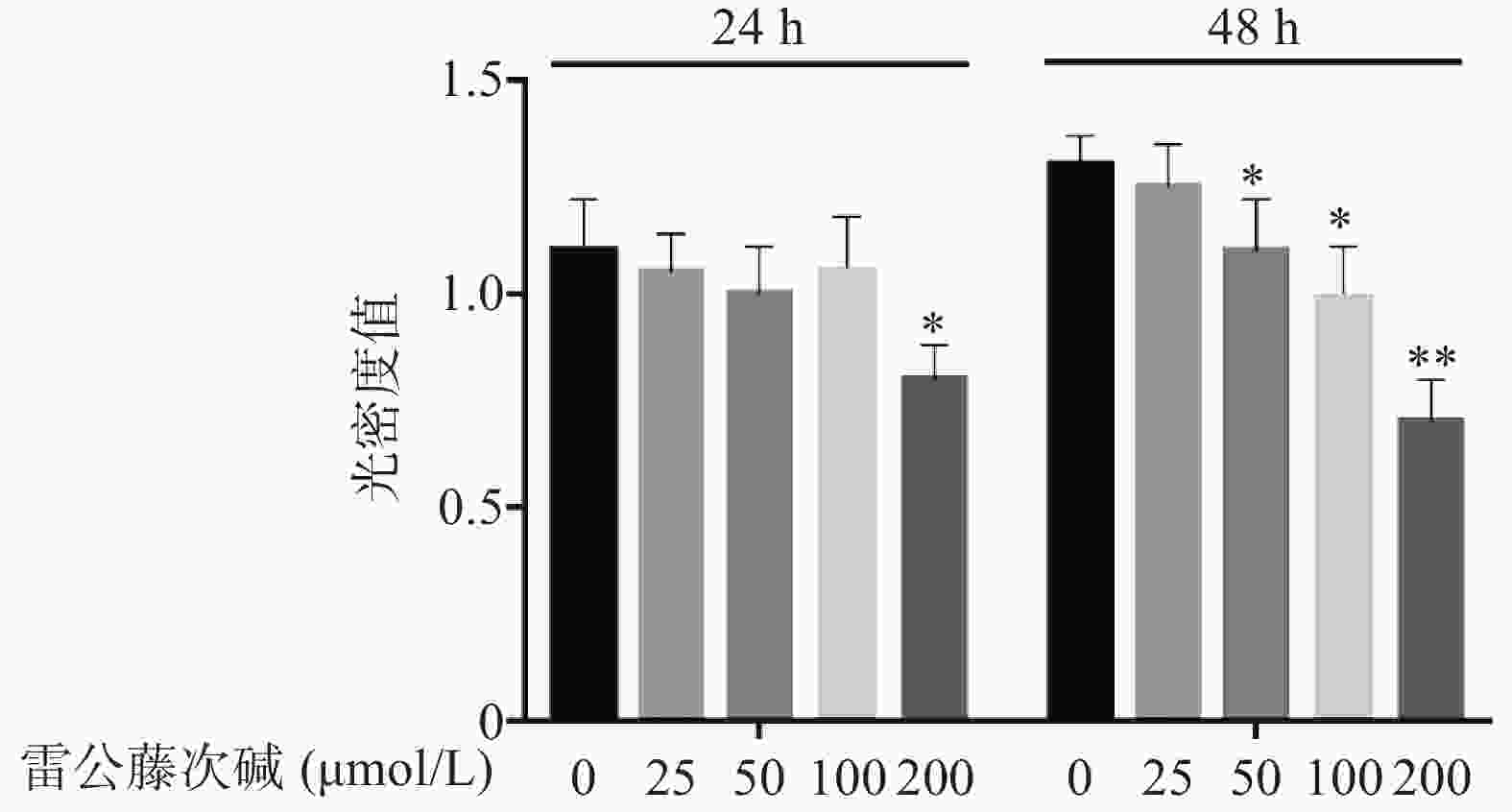
图 1 雷公藤次碱对RAW264.7细胞活力的影响
-
为了进一步研究雷公藤次碱体外抗炎活性,检测了雷公藤次碱对LPS刺激RAW264.7细胞分泌炎症因子的影响。结果显示(图2),给予LPS刺激后的RAW264.7细胞,NO、IL-1β、TNF-α和IL-6等促炎因子释放水平急剧增加(P<0.01),给予不同浓度雷公藤次碱(100、50和25 μmol/L)处理后,上述因子分泌水平呈剂量依赖性显著降低(P<0.01)。由此提示,雷公藤次碱的抗炎作用可能与抑制细胞释放NO、IL-1β、TNF-α和IL-6等促炎因子有关。
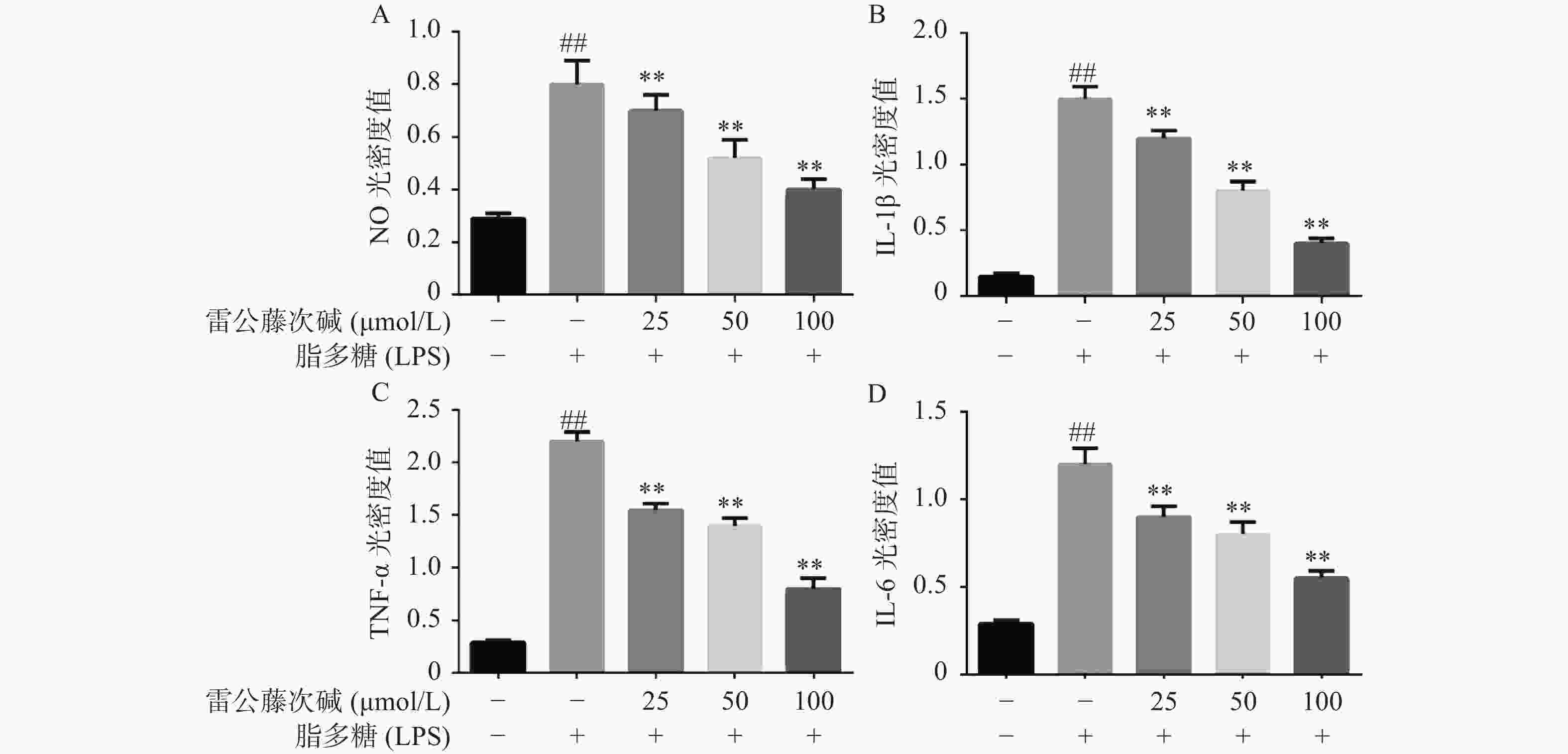
图 2 雷公藤次碱对LPS诱导RAW264.7细胞分泌炎症因子的影响
-
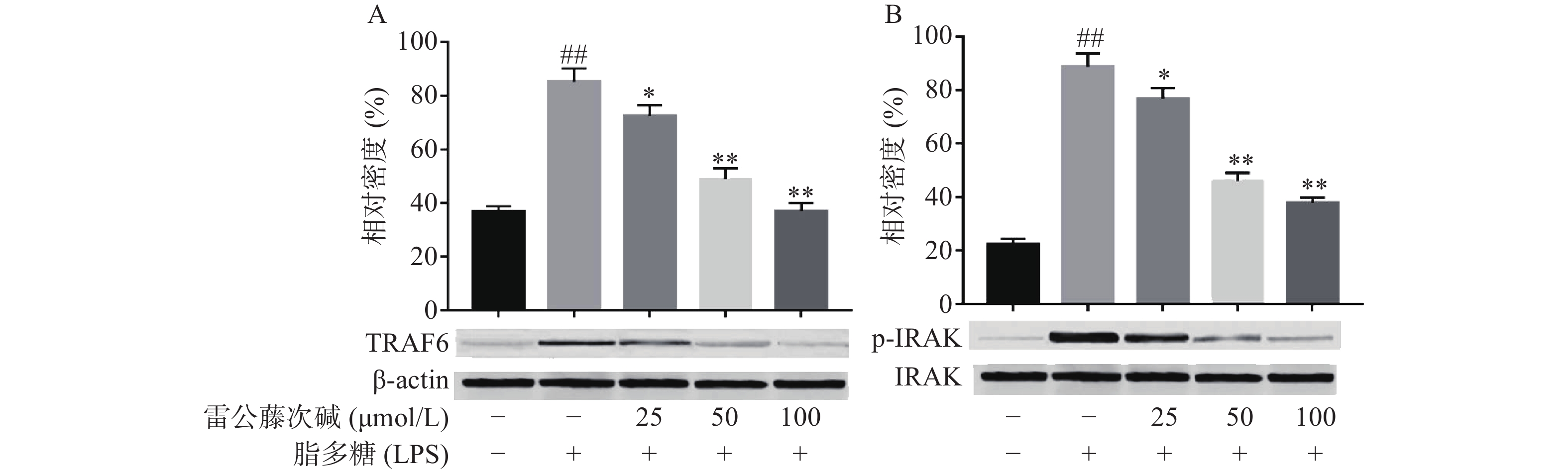
LPS是引发炎症的早期关键因素,可诱发多种细胞内信号事件,LPS-TLR4/MyD88信号通路可调节促炎基因表达,并控制炎症相关细胞因子的表达,其中IRAK及TRAF6是MyD88依赖性途径中的关键信号传导的分子。TLR4受体受LPS诱导后,可导致IRAK磷酸化从而脱离MyD88与TRAF6的结合体,使得游离的TRAF6活化后续信号转导途径[6]。为了评估雷公藤次碱对LPS激活TLR4/MyD88信号通路的影响,蛋白印迹法考察了雷公藤次碱对IRAK磷酸化和TRAF6表达的影响。结果显示(图3),与空白组相比,LPS刺激的模型组IRAK磷酸化和TRAF6表达明显增加(P<0.01);与模型组相比,雷公藤次碱各剂量组可显著抑制IRAK磷酸化和降低TRAF6表达(P<0.01)。
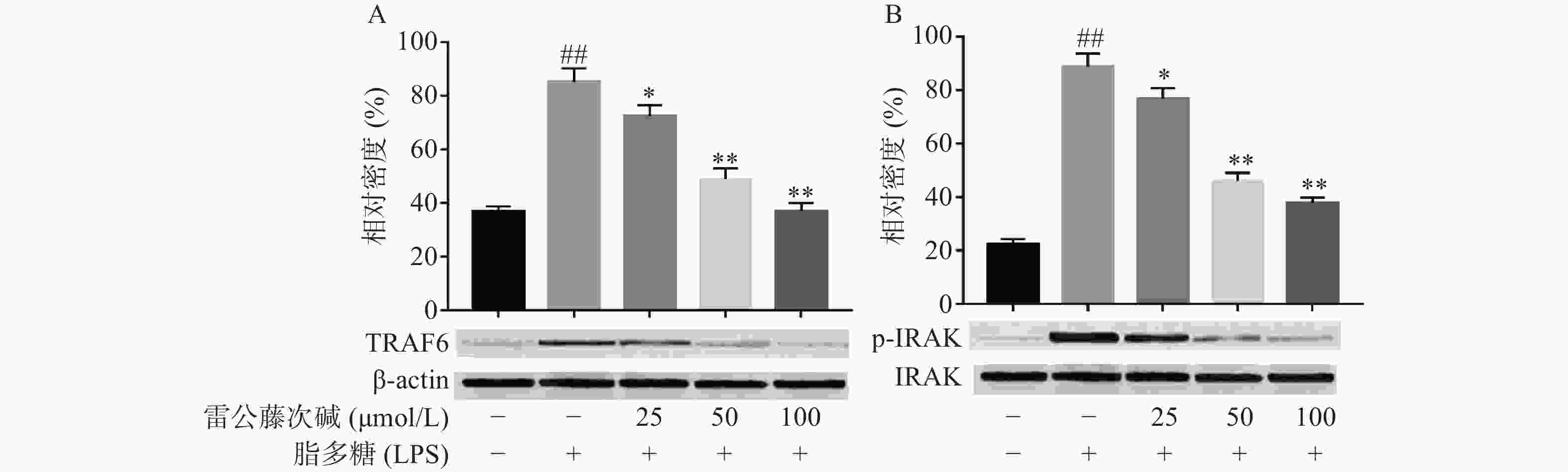
图 3 雷公藤次碱对TRAF6表达和IRAK磷酸化的表达的影响
-
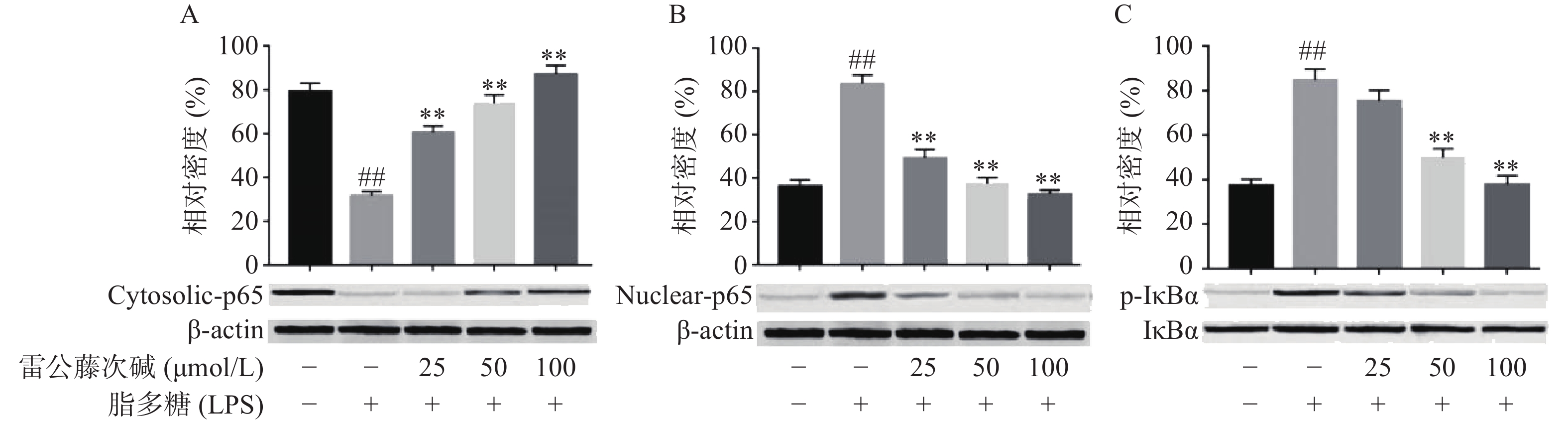
NF-κB是MyD88信号通路中重要的转录因子[7]。我们考察了雷公藤次碱对LPS诱导的RAW264.7细胞中NF-κB活化的潜在影响。如图4所示,RAW264.7细胞LPS刺激后,细胞核内NF-κB p65水平增高而细胞质内NF-κB p65水平降低,说明NF-κB p65活化后由细胞质进入到细胞核内,而给予雷公藤次碱处理后,细胞质内NF-κB p65水平增高而细胞核内NF-κB p65水平降低,说明雷公藤次碱以浓度依赖的方式显著地阻止了NF-κB p65从细胞质转运到细胞核。 NF-κB p65易位进入核内与TLR4途径中的IκBα磷酸化有关。因此,我们考察了雷公藤次碱对NF-κB抑制蛋白IκBα磷酸化的影响。结果表明,雷公藤次碱以浓度依赖的方式显著抑制LPS诱导的IκBα磷酸化和降解。
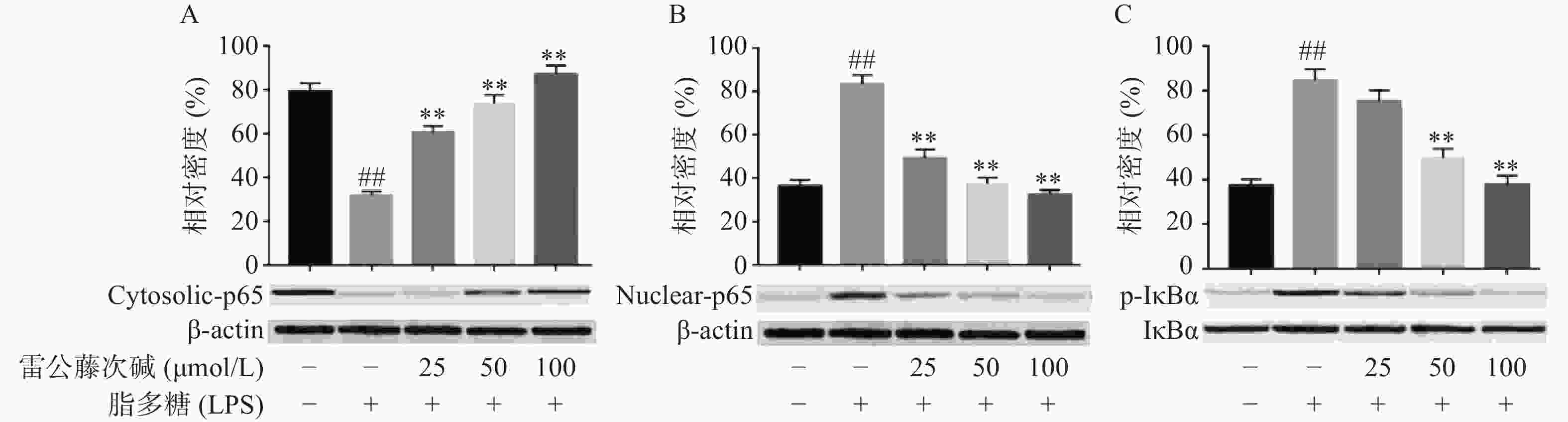
图 4 雷公藤次碱对IκBα磷酸化和NF-κB核转位的影响
-
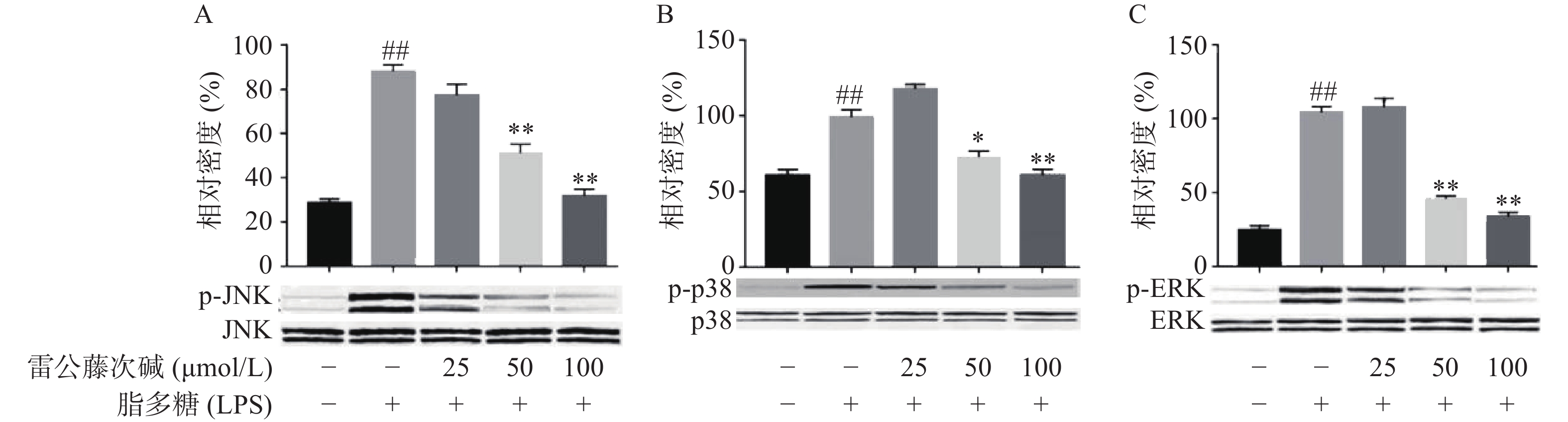
MAPKs是信号从细胞表面传导到细胞核内部的重要传递者。为了进一步研究雷公藤次碱是否调节MAPKs,考察了其对LPS诱导的RAW264.7细胞中ERK、p38和JNK等MAPKs磷酸化的影响。如图5所示,LPS暴露可显著促进RAW264.7细胞中ERK、JNK和p38的磷酸化。给予雷公藤次碱可以显著抑制LPS诱导的ERK、p38和JNK的磷酸化,但并未影响ERK、p38和JNK的表达。上述结果表明,MAPKs的磷酸化可能参与了雷公藤次碱对RAW264.7细胞中LPS刺激炎症的抑制作用。
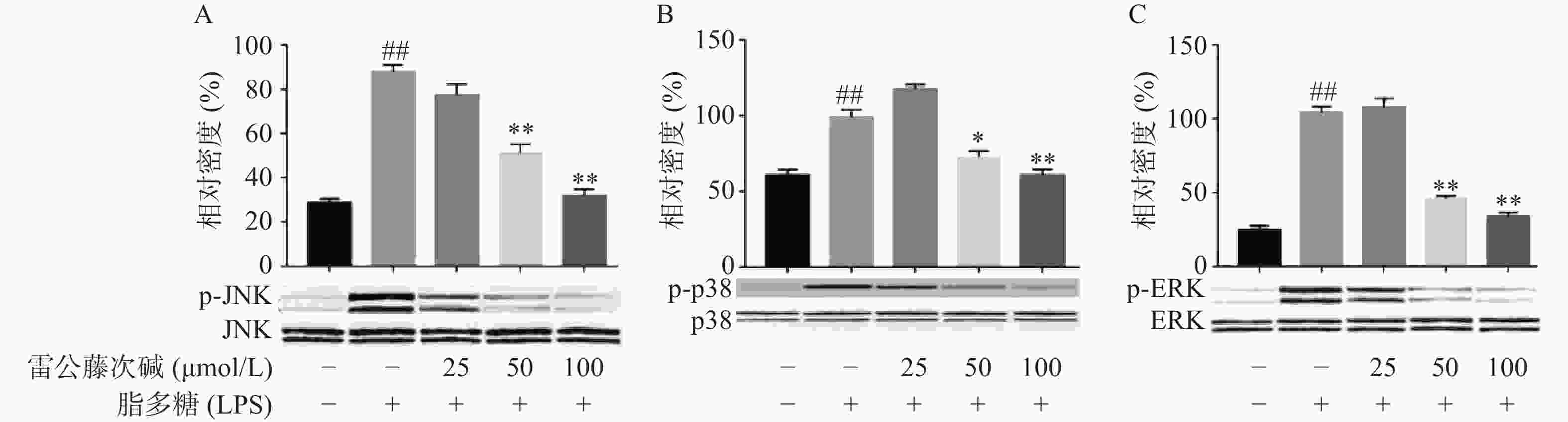
图 5 雷公藤次碱对p38, ERK和JNK MAPKs磷酸化的影响
-
雷公藤次碱是雷公藤的代表性生物碱成分之一,但是自从被分离以来,其生物学活性除了杀虫外,其他方面了解甚少[8]。本研究显示,雷公藤次碱对LPS诱导的RAW264.7细胞炎症模型显示出显著的抗炎活性。雷公藤次碱可减少LPS刺激引起的NO、IL-1β、TNF-α及IL-6等促炎因子的产生,并抑制LPS诱导的TLR4/MyD88通路中信号传导的分子IRAK磷酸化及TRAF6表达,从而影响MAPKs和NF-κB的激活,如图6所示。
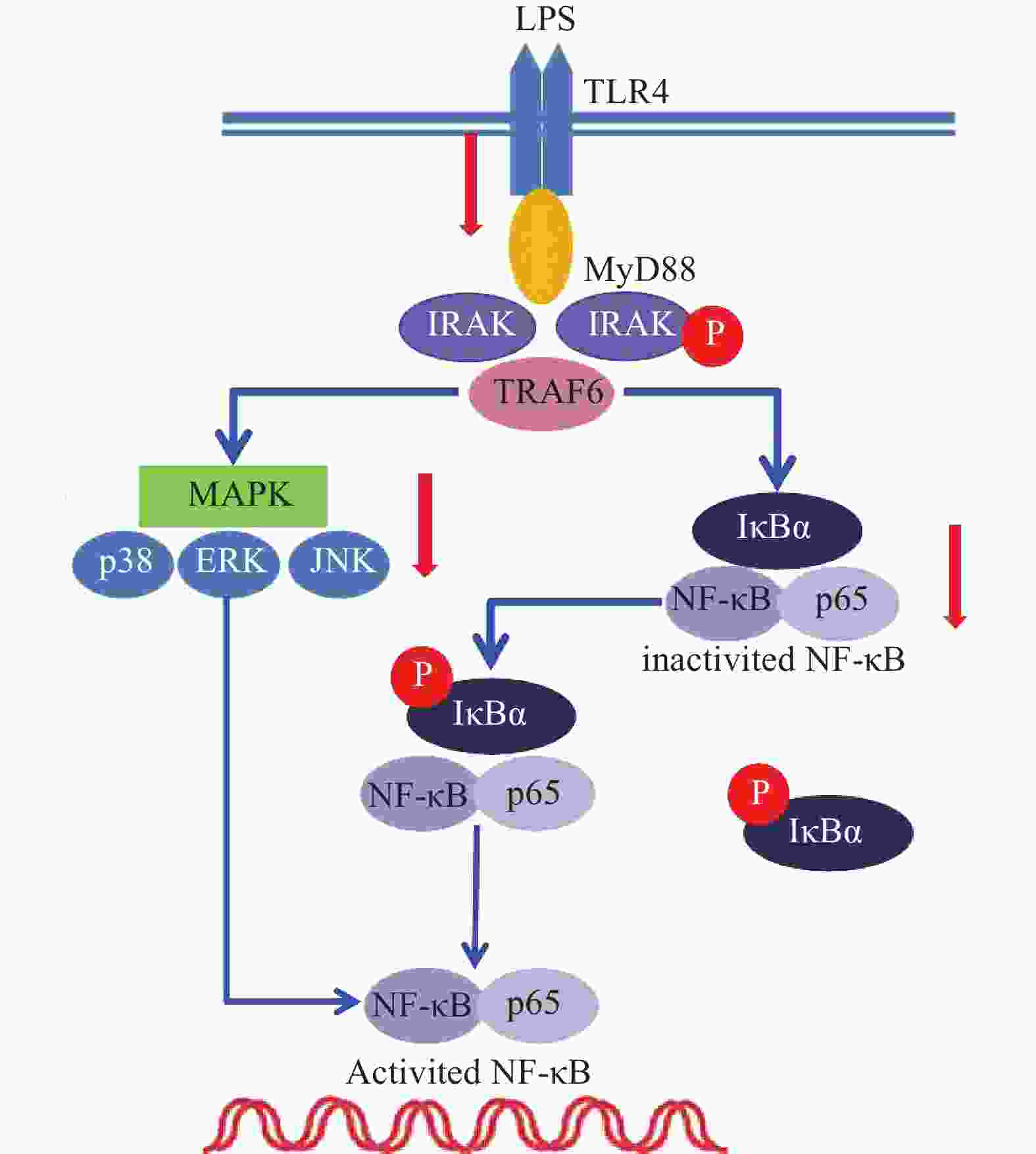
图 6 雷公藤次碱抗炎调控机制
已有证据显示,LPS可引发多种细胞内信号事件,包括导致TLR4,NF-κB及p38,ERK和JNK等丝裂素活化蛋白激酶(MAPKs)途径激活[9-10]。研究发现,LPS诱导后RAW264.7细胞Toll样受体4(TLR4)的表达明显增高,说明LPS是或可引导TLR4的配体与TLR4结合,二者结合后可启动细胞内一系列信号级联反应,最终诱导目的基因的表达。TLRs亚型中除TLR3外,均需要髓样分化因子88(MyD88)来激活下游释放信号分子的通路,这种途径成为MyD88依赖性途径。静息状态下,MyD88与IRAK及TRAF6形成细胞信号转导复合物,受到诱导后导致IRAK磷酸化,IRAK磷酸化后TRAF6从信号转导复合物中解离出来,TRAF6的活化可引起两条不同的信号转导途径,一条为MAPK家族(包括p38、JNK),另一条为活化MP3K(mitogen activated protein kinase kinase kinase)家族成员NIK(NF-κB inducing kinase),后者的磷酸化激活IκB激酶,导致IκB的磷酸化而使NF-κB/IκB复合物解离,NF-κB由此活化转位进入细胞核[11]。在本研究中,数据显示雷公藤次碱可以抑制IRAK磷酸化和TRAF6的表达。
NF-κB是一种多效性的转录因子,参与调控炎症反应、免疫、肿瘤等相关的细胞因子、趋化因子、黏附分子、炎症介质等的转录过程。IκB是其抑制蛋白,正常情况下二者结合而使NF-κB处于失活状态,IκBα是IκB的亚分子,受到LPS刺激,IκBα磷酸化增加,从而导致游离NF-κB p65释放并从细胞质转移至细胞核,与DNA上的相应位点结合,导致特定靶基因(如TNF-α、IL-1β和IL-6)的转录表达[12-13]。在本研究中,数据显示雷公藤次碱可抑制IκBα磷酸化,并以浓度依赖的方式阻止NF-κB p65由细胞质进入细胞核,从而阻止炎症相关因子的转录表达。
此外,NF-κB的易位也受ERK、p38和JNK等丝裂素活化蛋白激酶途径调节,该途径可控制炎症反应过程中细胞因子的合成和释放。丝裂素活化蛋白激酶激活后,细胞质或细胞核中的转录因子又被激活,从而触发引起生物学反应的靶基因的表达,包括促炎性介质的表达。本研究显示,LPS诱导的RAW264.7细胞中,雷公藤次碱可显著抑制ERK、JNK和p38的磷酸化。
Wilforine inhibits LPS-induced inflammatory response in RAW264.7 cells by regulating the TLR4/MyD88/TRAF6 signaling pathway
-
摘要:
目的 探讨雷公藤次碱抗炎活性及其作用机制。 方法 用细胞计数盒-8 (CCK-8)法考察雷公藤次碱对小鼠单核巨噬细胞白血病细胞RAW 264.7 增殖活性的影响,用酶联免疫吸附法(ELISA)检测雷公藤次碱对脂多糖(LPS)诱导的RAW264.7细胞分泌细胞因子一氧化氮(NO)、白细胞介素-1β(IL-1β)、 肿瘤坏死因子α(TNF-α)和白细胞介素6(IL-6)的影响,用免疫印迹法考察雷公藤次碱对白介素受体相关激酶(IRAK)、肿瘤坏死因子受体相关蛋白6(TRAF6)、核因子κB抑制因子α(IκBα)、核因子κB(NF-κB p65)、分裂原激活的蛋白激酶p38(p38)、c-Jun氨基末端激酶(JNK)、细胞外调节蛋白激酶(ERK)在LPS刺激的RAW264.7细胞中的表达及其磷酸化的影响。 结果 雷公藤次碱在25、50、100 μmol/L浓度下对RAW264.7细胞无显著毒性,并可显著抑制细胞因子NO、IL-1β、TNF-α和IL-6含量。免疫印迹法检测结果显示雷公藤次碱可显著抑制IRAK及TRAF6的表达;显著抑制ERK、P38和JNK的磷酸化;抑制IκBα的降解,降低NF-κB p65的核转运水平。 结论 雷公藤次碱具有体外抗炎活性,其作用机制可能介导TLR4/MyD88/TRAF6信号通路。 Abstract:Objective To investigate the anti-inflammatory effect and mechanism of wilforine. Methods Anti-inflammatory activity of wilforine was investigated in LPS-induced RAW264.7 cells. The cytokines production of RAW264.7 cells was analyzed by ELISA assay and the cell viability was assessed by CCK-8 method. The expression of TRAF6, the phosphorylation of IRAK, p38, ERK and JNK, the degradation of inhibitory κBα (IκBα) and the nuclear translocation of NF-κB p65 were further investigated by western blot. Results Triptolide had no significant toxicity to RAW264.7 cells at concentrations of 25, 50 and 100 μmol/L. and could significantly inhibit the contents of cytokines NO, IL-1β, TNF-α and IL-6. Wilforine significantly decreased the expression of TRAF6 and phosphorylation of IRAK, and inhibited the phosphorylation of ERK, p38, and JNK and degradation of IκBα, and reduced the level of nuclear translocation of NF-κB p65. Conclusion The anti-inflammatory activity of wilforine of LPS-induced RAW264.7 cells is probably via TLR4/MyD88/TRAF6 signaling pathway. -
Key words:
- Tripterygium wilfordii /
- wilforine /
- anti-inflammation /
- TLR4 /
- MyD88 /
- NF-κB
-
图 2 雷公藤次碱对LPS诱导RAW264.7细胞分泌炎症因子的影响
A. NO光密度值;B. IL-1β光密度值;C. TNF-α光密度值;D. IL-6光密度值*P<0.05,**P<0.01,与模型组比较;##P<0.01,与空白组比较
图 3 雷公藤次碱对TRAF6表达和IRAK磷酸化的表达的影响
A. TRAF6磷酸化;B. IRAK磷酸化*P<0.05,**P<0.01,与模型组比较;##P<0.01,与空白组比较
图 4 雷公藤次碱对IκBα磷酸化和NF-κB核转位的影响
A. 细胞质NF-κB p65含量;B. 细胞核NF-κB p65含量;C. IκBα磷酸化*P<0.05,**P<0.01,与模型组比较;##P<0.01,与空白组比较
图 5 雷公藤次碱对p38, ERK和JNK MAPKs磷酸化的影响
A. JNK磷酸化;B. p38磷酸化;C. ERK磷酸化*P<0.05,**P<0.01,与模型组比较;##P<0.01,与空白组比较
-
[1] 白雪, 付瑞嘉, 乐世俊, 等. 雷公藤治疗类风湿性关节炎研究进展[J]. 中草药, 2020, 51(1):265-275. doi: 10.7501/j.issn.0253-2670.2020.01.034 [2] 姚骥如, 孙莹, 罗顺葵, 等. 雷公藤多苷的临床应用进展[J]. 中国新药与临床杂志, 2010, 29(3):179-182. [3] 胡攀勇, 李振麟, 濮社班, 等. 雷公藤研究进展[J]. 中国野生植物资源, 2013, 32(2):1-3,7. doi: 10.3969/j.issn.1006-9690.2013.02.001 [4] ZHANG Y Q, XU W, LI H, et al. Therapeutic effects of total alkaloids of Tripterygium wilfordii Hook f. on collagen-induced arthritis in rats[J]. J Ethnopharmacol,2013,145(3):699-705. doi: 10.1016/j.jep.2012.11.018 [5] 刘莉, 闫君, 舒积成, 等. 雷公藤生物碱类成分及其药理活性研究进展[J]. 天然产物研究与开发, 2019, 31(12):2170-2181. doi: 10.16333/j.1001-6880.2019.12.022 [6] LI T, LI F, LIU X Y, et al. Synergistic anti-inflammatory effects of quercetin and catechin via inhibiting activation of TLR4-MyD88-mediated NF-κB and MAPK signaling pathways[J]. Phytother Res,2019,33(3):756-767. doi: 10.1002/ptr.6268 [7] 鲍璐璐, 崔立红. TLR4/MyD88/NF-κB信号通路的研究进展[J]. 胃肠病学和肝病学杂志, 2019, 28(5):568-572. doi: 10.3969/j.issn.1006-5709.2019.05.019 [8] 师宝君, 姬志勤, 张继文, 等. 昆明山海棠的杀虫活性及有效成分[J]. 昆虫学报, 2007, 50(8):795-800. doi: 10.3321/j.issn:0454-6296.2007.08.007 [9] LAI J L, LIU Y H, LIU C, et al. Indirubin inhibits LPS-induced inflammation via TLR4 abrogation mediated by the NF-kB and MAPK signaling pathways[J]. Inflammation,2017,40(1):1-12. doi: 10.1007/s10753-016-0447-7 [10] LIU F, SUN G Q, GAO H Y, et al. Angelicin regulates LPS-induced inflammation via inhibiting MAPK/NF-κB pathways[J]. J Surg Res,2013,185(1):300-309. doi: 10.1016/j.jss.2013.05.083 [11] 单佳铃, 程虹毓, 文乐, 等. TLR/MyD 88/NF-κB信号通路参与不同疾病作用机制研究进展[J]. 中国药理学通报, 2019, 35(4):451-455. doi: 10.3969/j.issn.1001-1978.2019.04.002 [12] 崔嵩, 刘学锋, 吴斌. NF-кB在肿瘤中的研究进展[J]. 现代肿瘤医学, 2009, 17(1): 134-137. 范慧婕, 谭章斌, 赵晓山, 等. 四逆散通过MAPKs/NF-κB途径保护脂多糖致Raw264.7的细胞炎症[J]. 暨南大学学报(自然科学与医学版), 2019, 40(1): 10-18. [13] KAMINSKA B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy: from molecular mechanisms to therapeutic benefits[J]. Biochim Biophys Acta,2005,1754(1-2):253-262. doi: 10.1016/j.bbapap.2005.08.017 -

 点击查看大图
点击查看大图
图(6)
计量
- 文章访问数: 17240
- HTML全文浏览量: 6892
- PDF下载量: 178
- 被引次数: 0

