

 下载:
下载:
-
烟碱型乙酰胆碱受体(nAChRs)属于一种配体门控离子通道蛋白,作用于突触间的快速信号传导,参与复杂的生理和病理过程。nAChRs是由α、β、γ、δ和ε等5个亚基构成的同源或异源五聚体,其生理功能取决于它的结构和分布。目前已发现的nAChRs亚基种类有α1-10、β1-4、δ、γ和ε共17种,分子量约300 kD,在突触前和突触后均有分布。突触前nAChRs增强神经递质活性,突触后nAChRs快速传导兴奋。而非突触性nAChRs在各种神经递质传递中发挥作用,可作为许多疾病的潜在药物靶点。
α7nAChR是nAChRs中的一个亚型,相对分子质量约56 kD,由5个α7亚基组成,每个亚基有502个氨基酸,亚基间会形成一个中心通道,亚基接合处有5个配体的结合位点[1]。其功能域包括有与配体结合位点的细胞外域(ECD)、调节细胞内信号传导的细胞内结构域(ICD)、形成离子孔和栅极的跨膜结构域(TMD)。在ECD和TMD的耦合区域还存在保守的半胱氨酸环[2],形成半胱氨酸环配体门控离子通道。α7nAChR具有高Ca2+通透性,是其他离子的20倍。
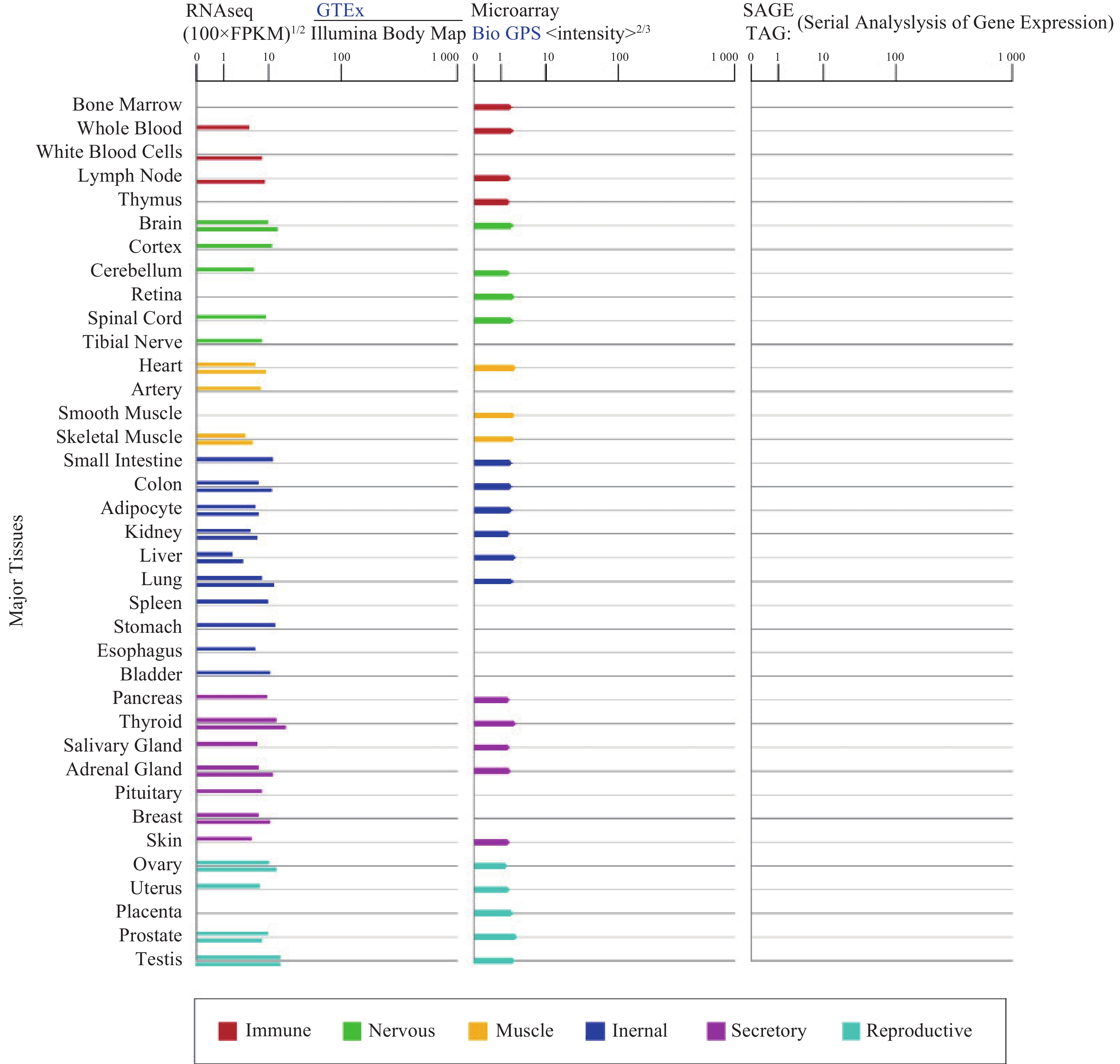
α7nAChR在人体各个组织器官广泛表达(图1),尤其在认知和记忆功能相关的海马和下丘脑高度表达,调节突触功能,影响神经递质释放[3]。α7nAChR在各种类型的免疫细胞如巨噬细胞[4]、淋巴细胞等中表达[5],是炎症治疗的关键受体;此外,α7nAChR也表达于非神经元细胞,像上皮细胞、血细胞、成纤维细胞、星形胶质细胞、小神经胶质细胞等[3, 6-7],与细胞凋亡和血管生成有关。
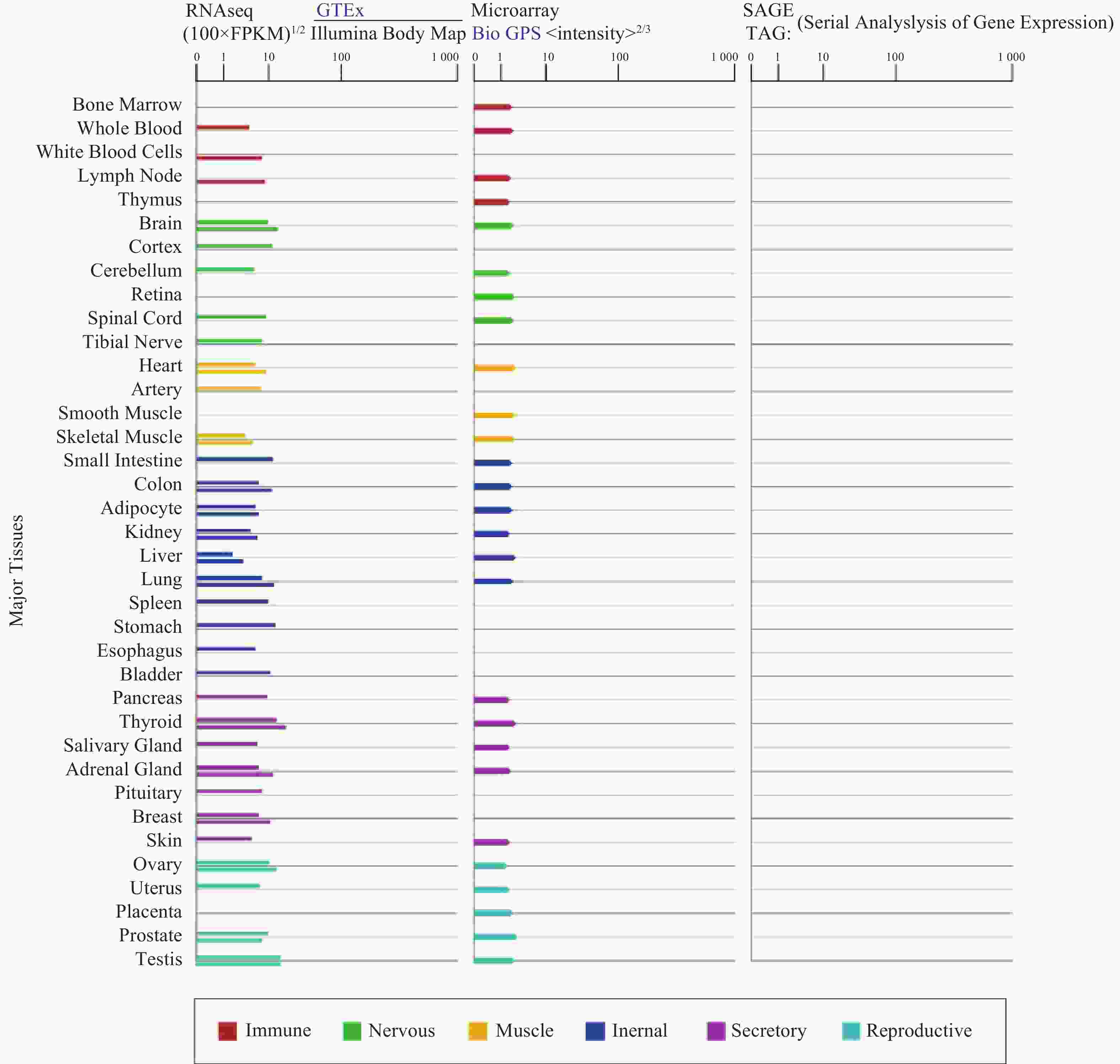
图 1 GTEx、Illumina、BioGPS和SAGE的CHRNA7基因在正常人体组织中的mRNA表达
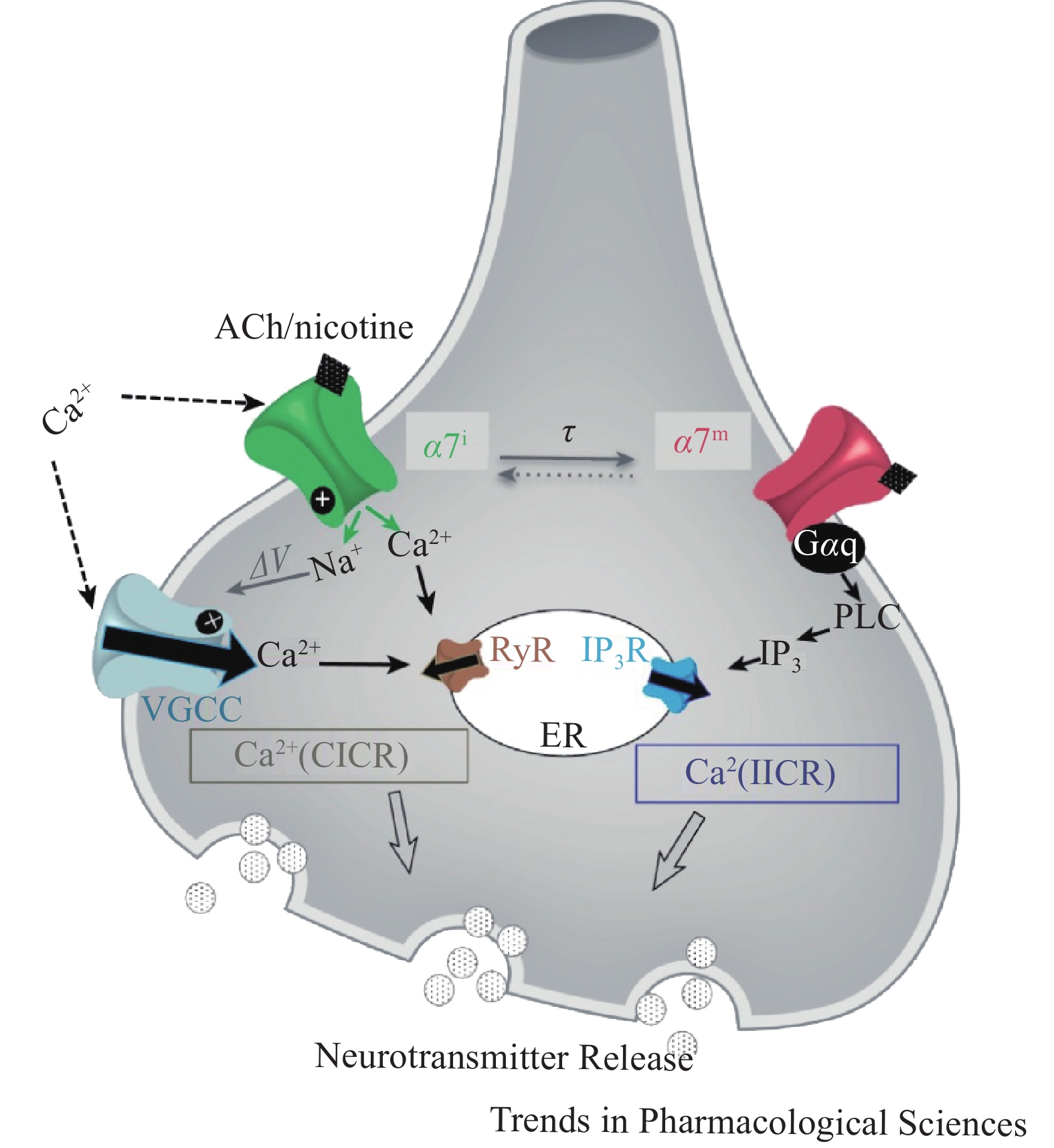
α7nAChR作为配体门控离子通道可以调节细胞内钙离子水平,表现出经典离子型通道和代谢型信号传导受体的双重功能,即钙离子诱导的钙释放(CICR)与异源三聚体GTP结合蛋白 (G蛋白)相关的肌醇三磷酸(IP3)诱导的钙释放(IICR)。α7nAChR介导的钙信号传导,可以通过开放离子通道Ca2+流入,诱发神经元兴奋,也可以直接与Gq蛋白偶联后由IP3介导Ca2+流入(图2)。蛋白质组学分析发现,在神经细胞、免疫细胞和上皮细胞中,有大量的蛋白质可能是nAChRs相互作用体的组成部分,为研究nAChRs信号传导和调节机制提供了新靶点。在啮齿动物的皮质、海马、纹状体等脑组织中已证实,nAChRs和G蛋白之间存在关联,但α7nAChR通过G 蛋白发出信号的机制尚不明确。α7nAChR的离子型和代谢型功能间的关系尚未阐明,α7nAChR代谢型受体信号传导可能辅助离子型通道快速离子电导[8]。

图 2 激动α7nAChRs后Na+和Ca2+流入示意图[8]
-
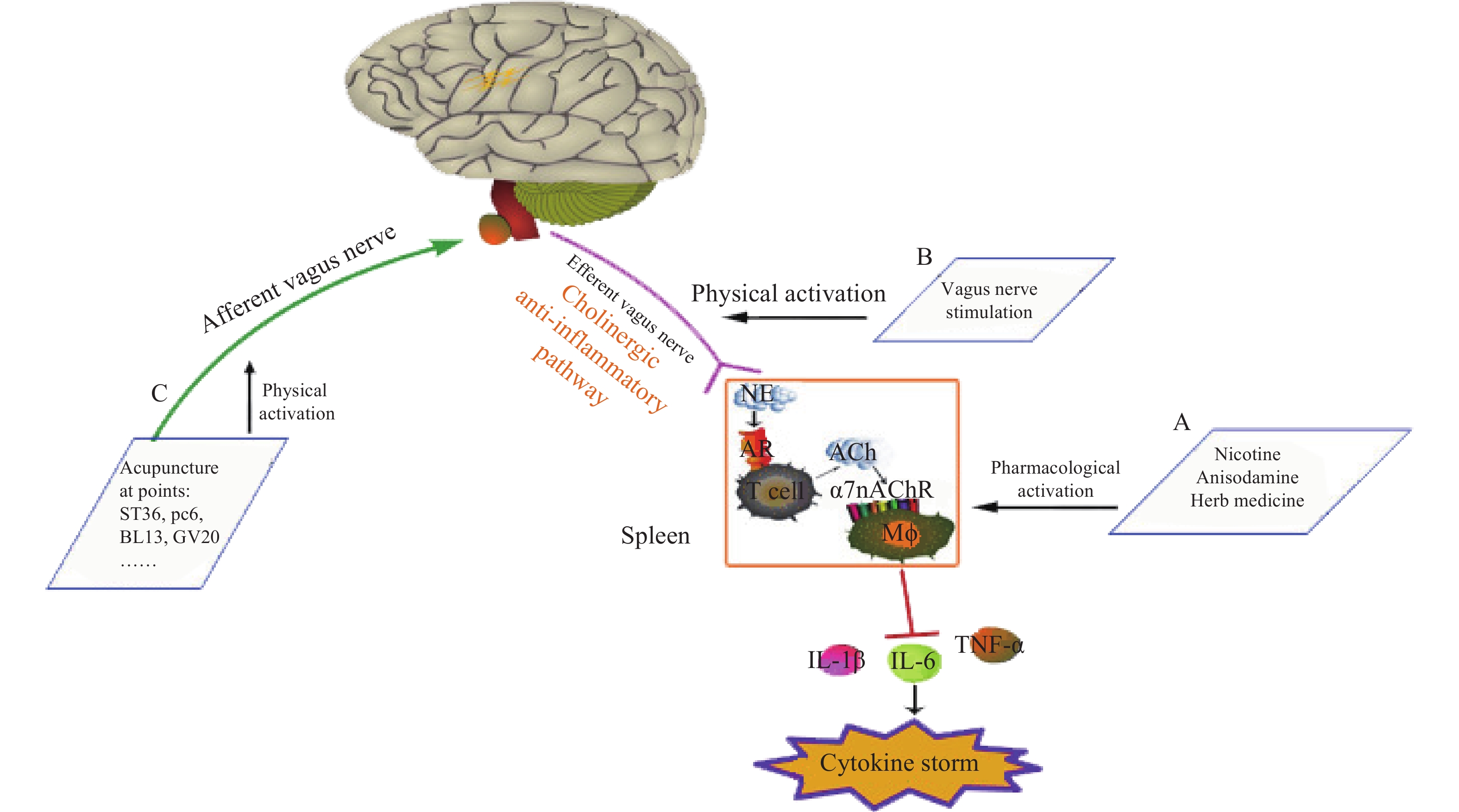
神经和免疫系统间存在复杂的相互作用,以维持健康稳态。Borovikov团队最早提出“胆碱能抗炎通路”(图3)概念[9]。迷走神经及其递质可抑制巨噬细胞活化。炎症反应时,炎症物质激活迷走传入神经,炎症信号传至延髓和下丘脑,迷走神经传入纤维会在延髓孤束核(NTS)处进行换元,信号直接传递到迷走神经运动背核(DMN)[1, 10],随后传出迷走神经,作用在支配脾脏的儿茶酚胺能纤维上,促进去甲肾上腺素(NE)释放。NE与脾脏乙酰胆碱转移酶(ChAT)+T细胞上的β2肾上腺素受体(β2AR)结合,释放的内源性乙酰胆碱(Ach)作用于巨噬细胞表面的α7nAChR,激活免疫细胞内的JAK2/STAT3信号通路,抑制了NF-κB的信号转导,炎症因子释放减少,发挥抗炎作用[11]。细胞内机制主要涉及抑制 NF-κB 的核转位,激活 JAK2/STAT3 级联反应,抑制线粒体上炎症小体的激活,从而抑制TNF、IL-1β等促炎细胞因子水平[12]。α7nAChR在胆碱能抗炎通路中是不可或缺的关键一环,例如,迷走神经电刺激(VNS)可降低小鼠体内TNF的水平,但在α7nAChR缺失小鼠体内,该效应消失[13]。
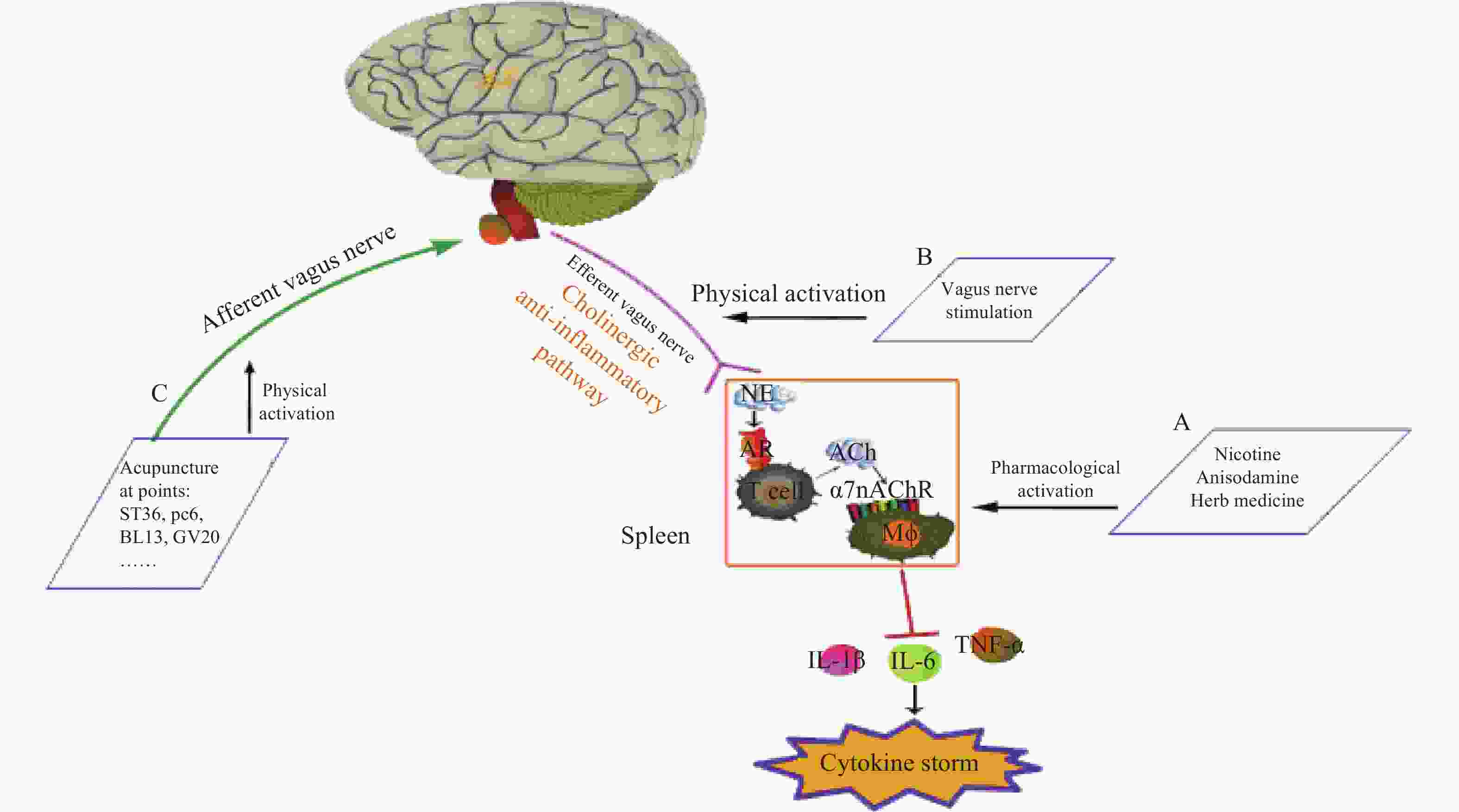
图 3 胆碱能抗炎通路[12]
迷走神经调节炎症的作用主要通过胆碱能抗炎通路[14]。与体液免疫的抗炎作用相比,胆碱能抗炎通路对炎症的响应更敏捷[15],它可以同时抑制多种促炎细胞因子,如 TNF、IL-1β等。刺激迷走神经或激活α7nAChR 能有效降低促炎细胞因子的生成,并提高动物在各种炎症性疾病中的存活率,如失血性休克、炎症性肠病(IBD),尤其是败血症[12],是神经-免疫反射的重要组成部分。据报道,在炎症相关疾病中,发现了迷走神经张力下降的现象,迷走神经张力降低可能是与发病机制相关的风险因素。因此,评估迷走神经张力并使之正常化可能是主要的治疗策略[16]。
-
炎症是人体对外部刺激的一种自主防御反应,但过度的炎症反应会攻击人体自身组织,导致广泛的组织损伤和功能障碍。机体调节炎症除体液免疫外,还有经典的神经-体液免疫抗炎通路:胆碱能抗炎通路。胆碱能抗炎通路在炎症反应早期便开始响应,且响应时间短,积极发挥早期抗炎作用,且由于迷走神经分布遍布全身脏器,能够灵活地进行全身或局部炎症反应调节[17],在目前炎症治疗中发挥不可替代的作用。
-
宿主因感染引发多器官功能损伤,使炎症因子(如IL-1、IL-6、TNF-α等)级联释放导致炎症失控,后期可能发展为严重的脓毒症和脓毒性休克,而脓毒症的病死率甚至超过25%,脓毒症如此高的致残致死率与免疫系统功能紊乱和炎症密不可分,调控炎症和免疫紊乱是治疗脓毒症的关键方法。
胆碱能抗炎通路通过对炎症的直接调节,能够实时并精准把控脓毒症的炎症反应进程,而α7nAChR是其中的关键调控靶点,在中枢和外周炎症治疗中都起着至关重要的作用[18]。电刺激脓毒症大鼠颈动脉窦神经能够明显改善脓毒症的炎症反应,迷走神经通过胆碱能抗炎通路参与并改善了大鼠体内的炎症反应[19],而切断大鼠的双侧颈迷走神经后,注射LPS后大鼠体内的炎症反应比假手术组更严重剧烈[9]。胆碱作为Ach水解的副产物,可以内源性非特异激动α7nAChR[20],若给予大鼠胆碱含量丰富的食物,能降低大鼠血清TNF水平,有效改善脓毒症的炎症反应[21],而将小鼠体内α7nAChR敲除后,再喂食胆碱含量丰富的食物后,血清内TNF水平无明显变化,无法改善脓毒症小鼠体内的严重炎症反应[22]。由此可知,目前迷走神经电刺激、α7nAChR受体激动剂(如PNU-282987、GTS-21、尼古丁等)[22]、胆碱酯酶抑制剂[23]等在脓毒症的治疗中极具前景。
-
创伤/失血性休克(T/HS)是一种遭受了外伤后严重出血,有效循环血量急剧降低引起的全身脏器组织血液灌注量不足的临床常见危重状态。T/HS的病死率高达 20%~40%,目前针对这一疾病的救治措施仍十分有限。但T/HS后会触发全身失控性炎性反应这一关键特点,为T/HS的防治提供了全新思路。激活迷走神经后的胆碱能抗炎通路,可在整体水平控制全身炎性反应[24],电刺激迷走神经后,肠道内CD103+树突状细胞调节Treg与 Th17的动态平衡,抑制T/HS后免疫细胞过度活化引起的全身性炎症风暴,减轻肺、心、肾等全身各器官损伤[25],以机体抗炎反射的各个环节为靶点,从整体水平控制全身炎性反应。电刺激迷走神经还会逆转失血性休克的低血压,降低失血性休克大鼠血浆的TNF-α水平,而迷走神经切断后静脉注射强效胆碱酯酶抑制剂THA,同样有抗炎抗休克作用,且肝脏中NF-κB p65水平明显下降,可以推测胆碱能抗炎通路是通过抑制NF-κB激活,降低促炎细胞因子转录合成,有效治疗休克。电刺激迷走神经前静脉注射α7nAChR拮抗剂α-BGT,抗炎和抗休克作用消失,表明α7nAChR在治疗T/HS中发挥关键作用[26]。
-
当组织一定时间内缺血后再次恢复血流(再灌注),组织的损伤程度会迅速增加,这称为缺血/再灌注损伤(I/R)。组织再灌注后,大量Ca2+内流,产生大量的自由基,攻击恢复血流后组织细胞,所以往往伴随着不受控制的炎症反应,尽管缺血再灌注损伤的致病机制非常的复杂,但炎症反应已被证实是其重要致病机制之一,TNF-α作为重要的促炎介质,通过中性粒细胞激活了机体内的炎症级联反应,促进炎症介质上调,加重组织的损伤。据报道,电针刺激足三里穴位后α7nAChR表达上调,减少炎性介质释放,减轻缺血再灌注中的肺损伤[27]。PNU 282987激活α7nAChRs后改善了I/R诱导的心肌自噬功能障碍,可能通过降低炎症反应、促进血管生成等方面保护心肌[28]。激活小胶质细胞中的α7nAChR,大脑I/R 损伤后的炎症与神经损伤均被改善[29],说明α7nAChR激动剂可能降低了I/R脑损伤中的炎症,发挥神经保护作用。金雀异黄素-3'-磺酸钠(GSS)通过α7nAChR-JAK2/STAT3信号通路抑制促炎细胞因子释放,阻断了神经炎症,改善神经功能,减少脑梗塞体积,从而减轻大鼠脑缺血损伤[30]。
最近,越来越多的临床前研究发现激活迷走神经可以保护缺血再灌注器官损伤,在I/R 中α7nAChR激活的作用和机制的研究具有重要价值,靶向激动α7nAChR药物的研发可为器官I/R 损伤治疗提供新的可能。
-
IBD是一种肠道炎症性疾病,涉及回肠、直肠和结肠,包括溃疡性结肠炎(UC)和克罗恩病(CD),尽管具体的致病因素和发病机制尚不明确,但由于肠道黏膜免疫系统异常而导致的炎症是IBD重要的发病机制之一,迄今为止,IBD仍无法治愈,且发病率与死亡率很高,很多患者药物治疗后病情毫无起色,目前也没有用于持续治疗IBD的有效药物。IBD患者多伴有自主神经功能障碍,且迷走神经活性明显降低[31]。
激活α7nAChR通过AMPK-mTOR-p70S6K 信号通路,诱导巨噬细胞自噬激活,减少促炎细胞因子释放,显著改善2,4,6-三硝基苯磺酸 (TNBS) 诱导的结肠炎[32],而α7nAChR阻断后,IBD的改善作用消失[33]。据报道,低功率治疗性超声能够非入侵性激活胆碱能抗炎通路,通过激活脾脏迷走神经,调节中性粒细胞、巨噬细胞和相关炎症因子水平改善IBD临床症状,α7nAChR在此抗炎通路中起着不可替代的作用[34]。
-
尼古丁也是激活α7nAChR的一种非特异性激动剂,尼古丁早已与许多癌症治疗相关,在卵巢癌中,尼古丁通过抑制RAS-MAPK信号抑制人卵巢癌细胞SKOV3和 TOV112D细胞增殖,对卵巢癌的治疗有潜在研究价值[35]。
有研究表明,迷走神经在各种癌症疾病中对机体有潜在保护作用,临床研究发现迷走神经切断后,患胃癌、结直肠癌、胆道癌和肺癌的风险增加[36-37],肺癌的死亡率也会增加[37]。在各种动物模型中,迷走神经切断后胰腺癌[38]和胃癌发病率明显上升[39-40],而激活迷走神经后,小鼠的肺癌和肝癌细胞转移数量明显减少[41],这可能与迷走神经激活后抑制TNF-α分泌,增强树突状细胞活性有关[42]。
-
脑卒中是一种急性脑血管疾病,由于脑内血管突然破裂或阻塞,血液无法流进大脑,脑组织将严重受损,脑卒中分为缺血性卒中和出血性卒中。在缺血性卒中中,脑组织灌注突然下降会引起兴奋性毒性、细胞大量坏死与凋亡,并发生剧烈的炎症反应,脑损伤后,中枢神经系统发出强烈的炎症信号,外周免疫细胞浸润并进一步加强局部炎症反应。例如,在缺血性卒中中,单核细胞/巨噬细胞及中性粒细胞是最先到达脑部损伤位置的细胞,其次是T淋巴细胞与B淋巴细胞,胆碱能系统还能调节星形胶质细胞和小胶质细胞对脑损伤的反应,因此,免疫反应调节是脑卒中疾病治疗的重要手段。ACh与小胶质细胞上的α7nAChR结合能下调促炎细胞因子,小胶质细胞与星形胶质细胞的抗炎通路均由 α7nAChR 介导。给予尼古丁后,星形胶质细胞中IL-1β、IL-6和TNFα均下调,星形胶质细胞的抗炎作用与抑制 NF-κB 和激活 Nrf2 信号传导有关[43]。中脑运动区(MLR)的高频刺激可通过影响胆碱能抗炎途径减轻大鼠的神经炎症,减少光血栓性脑卒中(PTS)后的病灶周围炎症,使IFN-γ、TNF-α和IL-1α的数量减少[44]。胆碱能信号通路调节神经和外周免疫的功能,在脑卒中治疗中有潜在前景。
-
类风湿性关节炎(RA)是一种自身免疫性疾病,以炎性滑膜炎为主,病因至今不明,多关节的炎症甚至会导致关节的畸形或丧失功能。α7nAChR被激活后通过影响免疫细胞的活性降低炎症反应,在RA小鼠模型中,GTS-21通过减少树突状细胞浸润,降低血清TNF-α和IL-6的水平,有效改善炎症。尼古丁激活α7nAChR后,通过胆碱能抗炎通路抑制了Th1和Th17细胞分化和巨噬细胞的迁移,调节Th1/Th2,促进Th2分化[45-48],抑制TNF-α表达[49],减轻关节炎。青藤碱(SIN)作为国内治疗RA的临床药物,在佐剂性关节炎大鼠模型和成纤维细胞样滑膜细胞上均发现,SIN通过α7nAChR 抑制 NF-κB 通路的激活治疗RA[50]。
RA 患者的迷走神经活性明显降低,初步研究结果表明,无创经皮迷走神经电刺激可以减轻关节炎炎症,抑制促炎细胞因子的释放[51-52],侵入性迷走神经电刺激也可以抑制患者全血中 TNF-α水平[53],但具体机制仍需更多实验性的探索。这些研究结果都表明 α7nAChR和胆碱能抗炎通路在调节炎症反应,改善关节炎中起关键作用[54]。
-
近年来,新冠肺炎席卷全球,这是一种威胁人类健康的严重急性呼吸系统综合症(ARDS),SARS-CoV-2 是一种单链正链 RNA 病毒。越来越多临床证据显示COVID-19患者体内会出现严重的炎症风暴,这是免疫系统对感染做出的过度反应,而本身伴有糖尿病、心血管相关疾病、肥胖、慢性呼吸系统疾病及癌症等的患者,更易发展成严重的肺炎、ARDS或多器官衰竭,造成非常高的发病率与死亡率[55-57]。
在没有针对COVID-19特效药与疫苗出现之前,抗炎是治疗COVID-19当务之急,理论上使激活胆碱能抗炎通路治疗或减轻COVID-19成为可能。黄连解毒汤作为一种中药配方,1700多年来一直用于治疗败血症[58]。被认为是治疗COVID-19的有效药物,研究表明,黄连解毒汤可能影响ACh和胆碱水平,表明其抗炎作用可能通过胆碱能抗炎通路[59]。清肺排毒汤是国内应用最广泛的治疗COVID-19的方剂[60]。临床观察表明清肺排毒汤能显著改善COVID-19患者的临床症状,减少不良反应,提高治疗效果[12]。清肺排毒汤中的一些化合物已经有报道,是作用于胆碱能突触途径的靶标。例如麻黄碱可抑制乙酰胆碱脂酶(AChE)活性[61],调节nAChR[62],这些结果表明清肺排毒汤可以调节胆碱能抗炎通路治疗炎症。
GammaCore是一种手持式、独立的无创迷走神经刺激设备,直接接触颈部皮肤表面刺激迷走神经。最近,它被临床用于治疗两种与COVID-19相关的呼吸道症状患者,并显示出良好治疗效果[63]。
-
线粒体是细胞的能量工厂,其多孔外膜上有酶和受体,几种酶复合物和辅助因子嵌入线粒体内膜,构成线粒体呼吸链。当线粒体酶复合物的活性下降时,线粒体DNA受损。在中枢神经系统炎症中,线粒体DNA受损后细胞能量代谢水平下降;在非炎性神经退行性疾病中,ATP合成减少,导致细胞死亡或变性[64]。
物质代谢伴随能量代谢。早在1848年,Bernard就研究了肝脏自主神经支配葡萄糖代谢,将迷走神经切断后,肝脏葡萄糖的输出减少;迷走神经刺激后加速糖原合成,提高肝脏葡萄糖水平并维持葡萄糖稳态,具体机制仍未可知[65]。而越来越多的证据表明α7nAChR和迷走神经在能量稳态的中枢和外周机制中起关键作用,α7nAChR与肥胖和相关并发症的病理生理学相关。α7nAChR激活后,通过影响大脑中下丘脑神经肽、多种神经递质系统、边缘系统和旁边缘系统抑制食欲。尼古丁可以提高啮齿动物对胰岛素的敏感性,改善葡萄糖稳态和胰岛素信号传导[66],有益脂质和葡萄糖代谢。据报道,激活α7nAChR后可以减少甘油三酯/脂肪酸(TG/FA)底物循环,并增强脂肪组织中甘油三酯(TG)的摄取[67]。糖尿病和肥胖小鼠中迷走神经的活性降低,调节肝葡萄糖生成的迷走神经信号减弱,肝脏内的巨噬细胞Kupffer细胞活性失调,肝脏糖异生基因显著下调[68],而VNS激活α7nAChR后可以减轻肥胖引起的代谢紊乱,治疗脂肪肝[69]。脂肪组织中胆碱能信号传递功能障碍可能是导致肥胖的机制,因此,靶向调控α7nAChR或可成为治疗肥胖的新机制。
-
非酒精性脂肪肝(NAFLD)是全球最常见肝病之一。作为一种多系统疾病,NAFLD临床表现一般为代谢异常[70],包括高血糖、高血脂、肥胖、糖尿病等[71-73]。NAFLD的病发与胰岛素抵抗和肝脏脂肪堆积有关,特征是慢性肝损伤和纤维化。肝脏是人体代谢最重要的器官,所以NAFLD又被称为代谢异常综合征[74],具体发病机制尚不明确。
α7nAChR通过迷走神经在脑-肝交互中发挥重要作用。有研究报道胆碱能抗炎通路治疗NAFLD,脂肪肝中α7nAChR表达上调,选择性激动α7nAChR后明显改善NAFLD损伤,改善肝纤维化[67]。更重要的是,α7nAChR敲除后能量消耗明显降低,体重增加,高血糖和高胰岛素症状消失[75],表明α7nAChR可能在能量稳态中发挥有益作用。
-
阿尔兹海默症(AD)临床初期主要表现为思维能力下降,近期记忆丢失并伴有认知障碍,后逐渐发展为记忆缺失,典型病理标志有细胞外淀粉样斑块、细胞内神经元纤维缠结、神经突营养不良等[76]。患者脑血流量下降[77]直接影响能量代谢,脑葡萄糖代谢损伤会降低乙酰辅酶A浓度,进而降低乙酰胆碱合成速率[78-79],影响胆碱能系统[80],自由基增多、细胞内Ca2+稳态失调使线粒体功能紊乱[81]。能量代谢下降和氧化应激又会增加AChE活性。脑能量代谢异常可能是AD重要的发病机制[82]。Tota等发现,氧化应激和葡萄糖利用降低使ACh mRNA上调,乙酰胆碱活性增强。水飞蓟宾通过改善脑能量代谢和胆碱能系统治疗AD认知功能下降[78]。
α7nAChR和能量代谢与AD的治疗紧密关联。α7nAChR激动剂PNU-282987处理原代小胶质细胞,线粒体的质量和耗氧量明显增加,通过α7nAChR调控小胶质细胞线粒体的机制通路,可能成为有效治疗AD的新途径[83]。据报道,α7nAChR表达下调时小鼠出现记忆障碍,同时ATP水平降低,很可能与氧化应激水平增加有关[78]。鼻内胰岛素可以预防链脲霉素诱导的乙酰胆碱活性下降、α7nAChR表达下调、血脑流量减少和线粒体功能衰退[84]。鼻内胰岛素增加ACh活性和α7nAChR表达可能是因为改善了氧化应激,提高了ATP水平[85]。
尽管在神经退行性疾病致病机制的研究仍有空白领域,但能确定的是胆碱能系统与能量代谢起着至关重要的作用,尤其α7nAChR与脑能量代谢息息相关,是治疗阿尔兹海默症的关键受体。
-
α7nAChR作为连接神经系统和免疫系统的关键纽带,在调节机体稳态中不可替代。很多研究报道,作用于α7nAChR可以改善能量代谢,尤其是线粒体功能,治疗非酒精脂肪肝、促进血管新生、保护心脏等。同时α7nAChR在神经退行性疾病方面与能量代谢紧密关联。目前通过治疗炎症,增强能量代谢,改善线粒体的功能,成为治疗神经退行性疾病的热门方向。
综上,α7nAChR在能量代谢中发挥不可替代的作用,是能量代谢相关疾病治疗的潜在靶点,作为胆碱能抗炎通路中的关键受体,研究前景广阔。
The latest research progress of α7 nicotinic acetylcholine receptor in cholinergic anti-inflammatory pathway
-
摘要: α7nAChR(α7 nicotinic acetylcholine receptor)属于烟碱型乙酰胆碱受体(nAChRs),是机体胆碱能抗炎通路的关键受体,在免疫系统的神经调控中发挥重要作用。近期研究发现,α7nAChR还参与调节免疫以外的多种生理病理过程,如非酒精性脂肪肝、血管新生、保护心脏等。此外,α7nAChR在神经退行性疾病等与能量代谢紧密关联。本文综述α7nAChR在治疗炎症、改善能量代谢中发挥的作用,以及α7nAChR参与治疗的新方向。
-
关键词:
- α7烟碱型乙酰胆碱受体 /
- 胆碱能抗炎通路 /
- 能量代谢 /
- 线粒体 /
- 神经退行性疾病
Abstract: α7 nicotinic acetylcholine receptor (α7nAChR) belongs to nicotinic acetylcholine receptors (nAChRs), which is the key receptor in cholinergic anti-inflammatory pathway and plays an important role in the neural regulation of immune system. Recent studies have found that α7nAChR is also involved in the regulation of various physiological and pathological processes other than immunity, such as non-alcoholic fatty liver, angiogenesis, heart protection, etc. In addition, α7nAChR is closely related to energy metabolism in neurodegenerative diseases. This article reviews the role of α7nAChR in treating inflammation and improving energy metabolism, and the new direction of α7nAChR involved in therapy. -
图 1 GTEx、Illumina、BioGPS和SAGE的CHRNA7基因在正常人体组织中的mRNA表达
(来源:https://www.genecards.org/cgi-bin/carddisp.pl?gene=CHRNA7&keywords=7nAChR)
-
[1] 王大伟, 周荣斌, 姚咏明. 胆碱能抗炎通路在炎症反应中的作用[J]. 生理科学进展, 2010, 41(3):217-220. [2] BOUZAT C, LASALA M, NIELSEN B E, et al. Molecular function of α7 nicotinic receptors as drug targets[J]. J Physiol,2018,596(10):1847-1861. doi: 10.1113/JP275101 [3] BERTRAND D, LEE C H, FLOOD D, et al. Therapeutic potential of α7 nicotinic acetylcholine receptors[J]. Pharmacol Rev, 2015, 67(4): 1025-1073. [4] MACKLIN K D, MAUS A D, PEREIRA E F, et al. Human vascular endothelial cells express functional nicotinic acetylcholine receptors[J]. J Pharmacol Exp Ther,1998,287(1):435-439. [5] CHEN L Y, LIU Z G, LI Y H, et al. Expression of neuronal acetylcholine receptor alpha 7 (nAChRalpha7) in peripheral blood CD(4)(+) T lymphocytes from asthmatic children[J]. Chin J Tuberc Respir Dis,2008,31(11):803-805. [6] KALKMAN H O, FEUERBACH D. Modulatory effects of α7 nAChRs on the immune system and its relevance for CNS disorders[J]. Cell Mol Life Sci,2016,73(13):2511-2530. doi: 10.1007/s00018-016-2175-4 [7] CORRADI J, BOUZAT C. Understanding the bases of function and modulation of α7 nicotinic receptors: implications for drug discovery[J]. Mol Pharmacol,2016,90(3):288-299. doi: 10.1124/mol.116.104240 [8] KABBANI N, NICHOLS R A. Beyond the channel: metabotropic signaling by nicotinic receptors[J]. Trends Pharmacol Sci,2018,39(4):354-366. doi: 10.1016/j.tips.2018.01.002 [9] BOROVIKOVA L V, IVANOVA S, ZHANG M, et al. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin[J]. Nature,2000,405(6785):458-462. doi: 10.1038/35013070 [10] 余江, 张恒, 张润峰. 胆碱能抗炎通路及其调控心脏炎症反应的作用[J]. 中国心血管病研究, 2021, 19(9):843-848. doi: 10.3969/j.issn.1672-5301.2021.09.017 [11] 马莉, 冷玉芳, 张梦婕, 等. 胆碱能抗炎通路对器官的保护研究进展[J]. 兰州大学学报(医学版), 2020, 46(1):10-15. doi: 10.13885/j.issn.1000-2812.2020.01.003 [12] QIN Z, XIANG K F, SU D F, et al. Activation of the cholinergic anti-inflammatory pathway as a novel therapeutic strategy for COVID-19[J]. Front Immunol,2020,11:595342. [13] WANG H, YU M, OCHANI M, et al. Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation[J]. Nature,2003,421(6921):384-388. doi: 10.1038/nature01339 [14] 陈卓, 何骁, 姚孟维, 等. 胆碱能炎症反射与炎症消退研究进展[J]. 中华烧伤杂志, 2021, 37(9):885-889. doi: 10.3760/cma.j.cn501120-20200609-00299 [15] 李祺, 刘霞. 胆碱能抗炎通路的信号机制研究[J]. 药学实践杂志, 2010, 28(5):325-327,338. [16] HOOVER D B. Cholinergic modulation of the immune system presents new approaches for treating inflammation[J]. Pharmacol Ther,2017,179:1-16. doi: 10.1016/j.pharmthera.2017.05.002 [17] 王娜, 苏跃. 全身炎症反应综合征治疗新策略: 胆碱能抗炎通路[J]. 医学与哲学, 2010, 31(22):42-43,77. [18] 资双凤, 李景辉, 刘蕾, 等. 胆碱能抗炎通路及其在脓毒症治疗中的作用[J]. 中南大学学报(医学版), 2020, 45(1):68-73. [19] BROGNARA F, CASTANIA J A, DIAS D P M, et al. Baroreflex stimulation attenuates central but not peripheral inflammation in conscious endotoxemic rats[J]. Brain Res,2018,1682:54-60. doi: 10.1016/j.brainres.2018.01.003 [20] SAPRONOV N S, NEZHINSKAYA G I, VLADYKIN A L. Effects of cholinergic agonists and antagonists under conditions of spleen denervation in rats with endotoxic shock[J]. Bull Exp Biol Med,2010,149(5):598-600. doi: 10.1007/s10517-010-1002-4 [21] RIVERA C A, WHEELER M D, ENOMOTO N, et al. A choline-rich diet improves survival in a rat model of endotoxin shock[J]. Am J Physiol,1998,275(4):G862-G867. [22] BENCHERIF M, LIPPIELLO P M, LUCAS R, et al. Alpha7 nicotinic receptors as novel therapeutic targets for inflammation-based diseases[J]. Cell Mol Life Sci,2011,68(6):931-949. doi: 10.1007/s00018-010-0525-1 [23] PAVLOV V A, PARRISH W R, ROSAS-BALLINA M, et al. Brain acetylcholinesterase activity controls systemic cytokine levels through the cholinergic anti-inflammatory pathway[J]. Brain Behav Immun,2009,23(1):41-45. doi: 10.1016/j.bbi.2008.06.011 [24] 郑紫磊, 郭睿, 郜思齐, 等. 迷走神经刺激活化胆碱能抗炎通路对创伤/失血性休克的干预作用[J]. 河北医药, 2021, 43(16):2531-2535. doi: 10.3969/j.issn.1002-7386.2021.16.031 [25] KOHOUTOVA M, HORAK J, JARKOVSKA D, et al. Vagus nerve stimulation attenuates multiple organ dysfunction in resuscitated porcine progressive Sepsis[J]. Crit Care Med,2019,47(6):e461-e469. doi: 10.1097/CCM.0000000000003714 [26] 李建国, 胡正芳, 杜朝晖, 等. 胆碱能抗炎通路对失血性休克大鼠保护作用的研究[J]. 中国危重病急救医学, 2005(1):24-27. [27] 宫丽荣, 董树安, 阚永星, 等. α7烟碱型乙酰胆碱受体在电针减轻肢体缺血再灌注兔肺损伤中的作用[J]. 中国中西医结合外科杂志, 2019, 25(4):443-447. doi: 10.3969/j.issn.1007-6948.2019.04.004 [28] 田甜, 何君艺, 樊素雄, 等. α7烟碱型乙酰胆碱受体对缺血再灌注损伤影响的研究现状[J]. 中国临床药理学杂志, 2020, 36(18):2914-2917. [29] MA Z, ZHANG Z L, BAI F H, et al. Electroacupuncture pretreatment alleviates cerebral ischemic injury through α7 nicotinic acetylcholine receptor-mediated phenotypic conversion of microglia[J]. Front Cell Neurosci,2019,13:537. [30] XIE J L, LI X, ZHANG L M, et al. Genistein-3'-sodium sulfonate ameliorates cerebral ischemia injuries by blocking neuroinflammation through the α7nAChR-JAK2/STAT3 signaling pathway in rats[J]. Phytomedicine,2021,93:153745. doi: 10.1016/j.phymed.2021.153745 [31] MUNYAKA P, RABBI M F, PAVLOV V A, et al. Central muscarinic cholinergic activation alters interaction between splenic dendritic cell and CD4+CD25- T cells in experimental colitis[J]. PLoS One,2014,9(10):e109272. doi: 10.1371/journal.pone.0109272 [32] SHAO B Z, WANG S L, FANG J, et al. Alpha7 nicotinic acetylcholine receptor alleviates inflammatory bowel disease through induction of AMPK-mTOR-p70S6K-mediated autophagy[J]. Inflammation,2019,42(5):1666-1679. doi: 10.1007/s10753-019-01027-9 [33] WAZEA S A, WADIE W, BAHGAT A K, et al. Galantamine anti-colitic effect: role of alpha-7 nicotinic acetylcholine receptor in modulating Jak/STAT3, NF-κB/HMGB1/RAGE and p-AKT/Bcl-2 pathways[J]. Sci Rep,2018,8(1):5110. doi: 10.1038/s41598-018-23359-6 [34] NUNES N S, CHANDRAN P, SUNDBY M, et al. Therapeutic ultrasound attenuates DSS-induced colitis through the cholinergic anti-inflammatory pathway[J]. EBioMedicine,2019,45:495-510. doi: 10.1016/j.ebiom.2019.06.033 [35] HARMYCH S J, KUMAR J, BOUNI M E, et al. Nicotine inhibits MAPK signaling and spheroid invasion in ovarian cancer cells[J]. Exp Cell Res,2020,394(1):112167. doi: 10.1016/j.yexcr.2020.112167 [36] OFFERHAUS G J, TERSMETTE A C, HUIBREGTSE K, et al. Mortality caused by stomach cancer after remote partial gastrectomy for benign conditions: 40 years of follow up of an Amsterdam cohort of 2633 postgastrectomy patients[J]. Gut,1988,29(11):1588-1590. doi: 10.1136/gut.29.11.1588 [37] AHSBERG K, OLSSON H, VON HOLSTEIN C S. Increased mortality in prostate carcinoma and smoking-related disease after parietal cell vagotomy: a long-term follow-up study[J]. Scand J Gastroenterol,2009,44(8):947-951. doi: 10.1080/00365520903039945 [38] OGAWA T, MAKINO T, MIZUMOTO K, et al. Promoting effect of truncal vagotomy on pancreatic carcinogenesis initiated with N-nitrosobis(2-oxopropyl)amine in Syrian golden hamsters[J]. Carcinogenesis,1991,12(7):1227-1230. doi: 10.1093/carcin/12.7.1227 [39] MORGENSTERN L. Vagotomy, gastroenterostomy and experimental gastic cancer[J]. Arch Surg,1968,96(6):920-923. doi: 10.1001/archsurg.1968.01330240066014 [40] KOWALEWSKI K. Relationship between vagotomy, peptic ulcer and gastric adenocarcinoma in rats fed 2, 7-diacetylaminogluorene[J]. Can J Surg,1973,16(3):210-217. [41] ERIN N, DUYMUŞ O, OZTÜRK S, et al. Activation of vagus nerve by semapimod alters substance P levels and decreases breast cancer metastasis[J]. Regul Pept,2012,179(1-3):101-108. doi: 10.1016/j.regpep.2012.08.001 [42] REIJMEN E, VANNUCCI L, DE COUCK M, et al. Therapeutic potential of the vagus nerve in cancer[J]. Immunol Lett,2018,202:38-43. doi: 10.1016/j.imlet.2018.07.006 [43] WINEK K, SOREQ H, MEISEL A. Regulators of cholinergic signaling in disorders of the central nervous system[J]. J Neurochem,2021,158(6):1425-1438. doi: 10.1111/jnc.15332 [44] SCHUHMANN M K, PAPP L, STOLL G, et al. Mesencephalic electrical stimulation reduces neuroinflammation after photothrombotic stroke in rats by targeting the cholinergic anti-inflammatory pathway[J]. Int J Mol Sci,2021,22(3):1254. doi: 10.3390/ijms22031254 [45] LIU D, LI T, LUO H, et al. The effect of the cholinergic anti-inflammatory pathway on collagen-induced arthritis involves the modulation of dendritic cell differentiation[J]. Arthritis Res Ther,2018,20(1):263. doi: 10.1186/s13075-018-1759-9 [46] LI S, ZHOU B, LIU B, et al. Activation of the cholinergic anti-inflammatory system by nicotine attenuates arthritis via suppression of macrophage migration[J]. Mol Med Rep,2016,14(6):5057-5064. doi: 10.3892/mmr.2016.5904 [47] WU S Y, LUO H, XIAO X Z, et al. Attenuation of collagen induced arthritis via suppression on Th17 response by activating cholinergic anti-inflammatory pathway with nicotine[J]. Eur J Pharmacol,2014,735:97-104. doi: 10.1016/j.ejphar.2014.04.019 [48] WU S Y, ZHAO H J, LUO H, et al. GTS-21, an α7-nicotinic acetylcholine receptor agonist, modulates Th1 differentiation in CD4+ T cells from patients with rheumatoid arthritis[J]. Exp Ther Med,2014,8(2):557-562. doi: 10.3892/etm.2014.1754 [49] VAN MAANEN M A, LEBRE M C, VAN DER POLL T, et al. Stimulation of nicotinic acetylcholine receptors attenuates collagen-induced arthritis in mice[J]. Arthritis Rheum,2009,60(1):114-122. doi: 10.1002/art.24177 [50] YI L, KE J Y, LIU J Y, et al. Sinomenine increases adenosine A 2A receptor and inhibits NF-κB to inhibit arthritis in adjuvant-induced-arthritis rats and fibroblast-like synoviocytes through α7nAChR[J]. J Leukoc Biol,2021,110(6):1113-1120. doi: 10.1002/JLB.3MA0121-024RRRR [51] KOOPMAN F A, SCHUURMAN P R, VERVOORDELDONK M J, et al. Vagus nerve stimulation: a new bioelectronics approach to treat rheumatoid arthritis? Best Pract Res Clin Rheumatol,2014,28(4):625-635. doi: 10.1016/j.berh.2014.10.015 [52] DREWES A M, BROCK C, RASMUSSEN S E, et al. Short-term transcutaneous non-invasive vagus nerve stimulation may reduce disease activity and pro-inflammatory cytokines in rheumatoid arthritis: results of a pilot study[J]. Scand J Rheumatol,2021,50(1):20-27. doi: 10.1080/03009742.2020.1764617 [53] COURTIES A, BERENBAUM F, SELLAM J. Vagus nerve stimulation in musculoskeletal diseases[J]. Joint Bone Spine,2021,88(3):105149. doi: 10.1016/j.jbspin.2021.105149 [54] LV J Q, JI X X, LI Z, et al. The role of the cholinergic anti-inflammatory pathway in autoimmune rheumatic diseases[J]. Scand J Immunol,2021,94(4):e13092. [55] ZHOU Y, CHI J W, LV W S, et al. Obesity and diabetes as high-risk factors for severe coronavirus disease 2019 (Covid-19)[J]. Diabetes Metab Res Rev,2021,37(2):e3377. [56] KWENANDAR F, JAPAR K V, DAMAY V, et al. Coronavirus disease 2019 and cardiovascular system: a narrative review[J]. Int J Cardiol Heart Vasc,2020,29:100557. [57] CHEN N S, ZHOU M, DONG X, et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study[J]. Lancet,2020,395(10223):507-513. doi: 10.1016/S0140-6736(20)30211-7 [58] WEI Y F, SHAN L, QIAO L M, et al. Protective effects of Huang-Lian-Jie-du-Tang against polymicrobial Sepsis induced by cecal ligation and puncture in rats[J]. Evid Based Complement Alternat Med,2013,2013:909624. [59] XU D Q, LV Y, WANG J S, et al. Deciphering the mechanism of Huang-Lian-Jie-Du-Decoction on the treatment of Sepsis by formula decomposition and metabolomics: enhancement of cholinergic pathways and inhibition of HMGB-1/TLR4/NF-κB signaling[J]. Pharmacol Res,2017,121:94-113. doi: 10.1016/j.phrs.2017.04.016 [60] REN J L, ZHANG A H, WANG X J. Traditional Chinese medicine for COVID-19 treatment[J]. Pharmacol Res,2020,155:104743. doi: 10.1016/j.phrs.2020.104743 [61] MAIZEL' E B, ROZENGART E V, KHAKIMOV I U P, et al. Ephedrine, salsoline and cytisine derivatives as substrates and inhibitirs of cholinesterases[J]. Biokhimiia,1978,43(7):1150-1156. [62] KINALI M, BEESON D, PITT M C, et al. Congenital myasthenic syndromes in childhood: diagnostic and management challenges[J]. J Neuroimmunol,2008,201-202:6-12. doi: 10.1016/j.jneuroim.2008.06.026 [63] STAATS P, GIANNAKOPOULOS G, BLAKE J, et al. The use of non-invasive vagus nerve stimulation to treat respiratory symptoms associated with COVID-19: a theoretical hypothesis and early clinical experience[J]. Neuromodulation,2020,23(6):784-788. doi: 10.1111/ner.13172 [64] LU F, SELAK M, O'CONNOR J, et al. Oxidative damage to mitochondrial DNA and activity of mitochondrial enzymes in chronic active lesions of multiple sclerosis[J]. J Neurol Sci,2000,177(2):95-103. doi: 10.1016/S0022-510X(00)00343-9 [65] PAVLOV V A, TRACEY K J. The vagus nerve and the inflammatory reflex: linking immunity and metabolism[J]. Nat Rev Endocrinol,2012,8(12):743-754. doi: 10.1038/nrendo.2012.189 [66] WU J, JIAO Z Y, LI R Z, et al. Cholinergic activation suppresses palmitate-induced macrophage activation and improves acylation stimulating protein resistance in co-cultured adipocytes[J]. Exp Biol Med (Maywood),2017,242(9):961-973. doi: 10.1177/1535370217700522 [67] NISHIO T, TAURA K, IWAISAKO K, et al. Hepatic vagus nerve regulates Kupffer cell activation via α7 nicotinic acetylcholine receptor in nonalcoholic steatohepatitis[J]. J Gastroenterol,2017,52(8):965-976. doi: 10.1007/s00535-016-1304-z [68] DENG J L, WANG M, GUO Y K, et al. Activation of α7nAChR via vagus nerve prevents obesity-induced insulin resistance via suppressing endoplasmic Reticulum stress-induced inflammation in Kupffer cells[J]. Med Hypotheses,2020,140:109671. doi: 10.1016/j.mehy.2020.109671 [69] YKI-JÄRVINEN H. Non-alcoholic fatty liver disease as a cause and a consequence of metabolic syndrome[J]. Lancet Diabetes Endocrinol,2014,2(11):901-910. doi: 10.1016/S2213-8587(14)70032-4 [70] KATSIKI N, MIKHAILIDIS D P, MANTZOROS C S. Non-alcoholic fatty liver disease and dyslipidemia: an update[J]. Metabolism,2016,65(8):1109-1123. doi: 10.1016/j.metabol.2016.05.003 [71] WOO BAIDAL J A, LAVINE J E. The intersection of nonalcoholic fatty liver disease and obesity[J]. Sci Transl Med,2016,8(323):323rv1. [72] BIRKENFELD A L, SHULMAN G I. Nonalcoholic fatty liver disease, hepatic insulin resistance, and type 2 diabetes[J]. Hepatology,2014,59(2):713-723. doi: 10.1002/hep.26672 [73] BUZZETTI E, PINZANI M, TSOCHATZIS E A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD)[J]. Metabolism,2016,65(8):1038-1048. doi: 10.1016/j.metabol.2015.12.012 [74] KIMURA K, INABA Y, WATANABE H, et al. Nicotinic alpha-7 acetylcholine receptor deficiency exacerbates hepatic inflammation and fibrosis in a mouse model of non-alcoholic steatohepatitis[J]. J Diabetes Investig,2019,10(3):659-666. doi: 10.1111/jdi.12964 [75] TING K K, BREW B, GUILLEMIN G. The involvement of astrocytes and kynurenine pathway in Alzheimer's disease[J]. Neurotox Res,2007,12(4):247-262. doi: 10.1007/BF03033908 [76] O'BRIEN J T, EAGGER S, SYED G M, et al. A study of regional cerebral blood flow and cognitive performance in Alzheimer's disease[J]. J Neurol Neurosurg Psychiatry, 1992, 55(12): 1182-1187. https://pubmed.ncbi.nlm.nih.gov/1479398/https://pubmed.ncbi.nlm.nih.gov/1479398/ [77] DE LA MONTE S M. Brain insulin resistance and deficiency as therapeutic targets in Alzheimer’s disease[J]. Curr Alzheimer Res,2012,9(1):35-66. doi: 10.2174/156720512799015037 [78] TOTA S, KAMAT P K, SHUKLA R, et al. Improvement of brain energy metabolism and cholinergic functions contributes to the beneficial effects of silibinin against streptozotocin induced memory impairment[J]. Behav Brain Res,2011,221(1):207-215. doi: 10.1016/j.bbr.2011.02.041 [79] HOYER S. Glucose metabolism and insulin receptor signal transduction in Alzheimer disease[J]. Eur J Pharmacol,2004,490(1-3):115-125. doi: 10.1016/j.ejphar.2004.02.049 [80] ZEEVALK G D, BERNARD L P, SINHA C, et al. Excitotoxicity and oxidative stress during inhibition of energy metabolism[J]. Dev Neurosci,1998,20(4-5):444-453. doi: 10.1159/000017342 [81] NAVARRO E, GONZALEZ-LAFUENTE L, PÉREZ-LIÉBANA I, et al. Heme-oxygenase I and PCG-1α regulate mitochondrial biogenesis via microglial activation of Alpha7 nicotinic acetylcholine receptors using PNU282987[J]. Antioxid Redox Signal,2017,27(2):93-105. doi: 10.1089/ars.2016.6698 [82] PLASCHKE K, HOYER S. Action of the diabetogenic drug streptozotocin on glycolytic and glycogenolytic metabolism in adult rat brain cortex and Hippocampus[J]. Int J Dev Neurosci,1993,11(4):477-483. doi: 10.1016/0736-5748(93)90021-5 [83] PATHAN A R, VISWANAD B, SONKUSARE S K, et al. Chronic administration of pioglitazone attenuates intrace-rebroventricular streptozotocin induced-memory impairment in rats[J]. Life Sci,2006,79(23):2209-2216. doi: 10.1016/j.lfs.2006.07.018 [84] RAJASEKAR N, NATH C, HANIF K, et al. Intranasal insulin improves cerebral blood flow, Nrf-2 expression and BDNF in STZ (ICV)-induced memory impaired rats[J]. Life Sci,2017,173:1-10. doi: 10.1016/j.lfs.2016.09.020 [85] RAJASEKAR N, DWIVEDI S, NATH C, et al. Protection of streptozotocin induced insulin receptor dysfunction, neuroinflammation and amyloidogenesis in astrocytes by insulin[J]. Neuropharmacology,2014,86:337-352. doi: 10.1016/j.neuropharm.2014.08.013 -
 点击查看大图
点击查看大图
图(3)
计量
- 文章访问数: 8881
- HTML全文浏览量: 4485
- PDF下载量: 120
- 被引次数: 0
