

 下载:
下载:

-
前列腺癌(prostate cancer,PCa)是全球范围内第二位最常见的男性癌症,也是全球第二大致死癌症[1]。据统计,我国前列腺癌的发病率呈逐年上升的趋势,其中,上海地区该病的发病率和病死率较高,但总体而言,亚洲国家前列腺癌的发病率明显低于西方国家[1-3]。在前列腺癌的治疗中,局限性前列腺癌常规行根治性前列腺切除术或放射治疗,而对于进展期患者则需要接受内分泌治疗即雄激素阻断治疗(androgen deprivation therapy,ADT)。虽然ADT在早期可以明显抑制激素依赖性前列腺癌的进展,但是随着治疗时间的延长,激素依赖性前列腺癌会逐渐失去对ADT的反应而转化成为去势抵抗性前列腺癌(castration-resistant prostate cancer, CRPC),目前对CRPC尚无有效的治疗方法。因此,亟需深入挖掘CRPC病程进展中的关键基因和蛋白,为CRPC提供潜在治疗靶标。
Abula等[4]进行了PCa蛋白质组学文献的数据挖掘,发现了前列腺肿瘤组织和正常组织中41种差异表达蛋白,并构建了蛋白相互作用网络。其中,3-甲基巴豆酰辅酶A羧化酶β亚基(3-methylcrotonyl-coenzyme A carboxylase β subunit,MCCB)就是其中的重要差异蛋白之一。MCCB是生物素依赖性羧化酶(3-methylcrotonyl-coenzyme A carboxylase,MCC)的两个组成亚基之一,由563个氨基酸组成[5-8],MCCC2是该蛋白的编码基因。已有研究报道,MCCC2是一种致癌基因,与包括肝细胞癌、结直肠癌以及乳腺癌在内的多种肿瘤的形成和发展密切相关[9-11],有望成为肿瘤治疗干预的潜在靶标。
在本研究中,我们主要探讨了MCCC2与前列腺癌的关系。首先,采用慢病毒与质粒构建了稳定低表达MCCC2的DU145细胞株并通过Western blot和qPCR分析敲减率;然后,系统考察敲减MCCC2对DU145前列腺癌细胞的增殖、迁移以及凋亡的影响,明确下调MCCC2与DU145细胞系肿瘤生物学特性的改变的相关性。
-
人前列腺癌DU145细胞,来源于海军军医大学附属长海医院泌尿外科,培养于含10%胎牛血清的RPMI 1640培养基中。
-
CCK-8试剂盒(上海翊圣生物科技有限公司);兔MCCC2多克隆抗体、GAPDH抗体以及HRP标记山羊抗兔二抗(美国CST公司);4%多聚甲醛固定液以及结晶紫染色液(碧云天生物技术有限公司);细胞凋亡试剂盒(南京凯基生物有限公司)。
-
DU145细胞复苏后接种于含10%灭活的胎牛血清,100 U/ml青霉素和100 μg/ml链霉素的DMEM培养液中,置于37 ℃,5% CO2细胞培养箱中生长,每隔3 d传代一次。将shRNA质粒复合物加到含有细胞和培养基的培养板的孔中,来回轻柔摇晃细胞培养皿。细胞在CO2培养箱中37 ℃孵育24 ~ 48 h进行转染;再用嘌呤霉素筛选低表达MCCC2-DU145的稳定细胞株。
-
将复苏的细胞接种于大的培养皿中。3 d后细胞以3×103个/孔,接种于96孔板。分别在0、24、48、72、96 h后,加CCK-8试剂,在450 nm处测定A值,根据A值分析结果。
-
取出DU145细胞、NC-DU145细胞以及shMCCC2-DU145细胞,弃掉上清,加入PBS缓冲液洗3次,每次1 ml;加入细胞裂解液(Western及IP细胞裂解液∶PMSF = 100∶1),吹打细胞,置冰上裂解20 min;收集细胞裂解液于1.5 ml的EP管中,4 ℃,12000 r/min,离心15 min;上清液即为提取的细胞总蛋白溶液,可用于BCA法定量;加入5 ×蛋白加样缓冲液,沸水中煮5 min,于−20 ℃保存,可用于后续的Western blot实验。
-
配胶,捡漏,加入样品,先将电压调为80 V,待marker条带分离后,将电压调制120 V,直到蛋白样品跑至凝胶底部,停止电泳;恒流250 mA,冰浴90 min的条件下转膜;染色,封闭,再孵育MCCC2抗体以及二抗,置于Tanon5200光密度扫描仪中进行拍照;用Image J 灰度分析差异性蛋白条带。
-
将复苏的细胞接种于培养皿中,待对数生长期时,细胞以5 × 105个/孔接种于6孔板中,在37 ℃,5% CO2的恒温培养箱中孵育;48 h后,将培养基吸掉,加入PBS洗涤3次,每次1 ml;加入RA2裂解液,每孔500 μl,置于冰上1 min;将裂解液收集于内套管中,在12000 r/min下离心1 min;取出,将外套管中的液体倒去,向内套管中加入500 μl洗液,在12000 r/min下离心1 min;重复上述步骤一次;重复上述步骤,但是这次不加洗液,在12000 r/min下离心1 min;取出,弃去外套管,将内套管移入新的1.5 ml Ep管中,加入30 μl的洗脱液,室温静置2 min,在12000 r/min下离心1 min;得到总的RNA,并测定其浓度。
-
按照1 000 μg逆转录,总反应体系为20 μl。反应条件为:37 ℃,15 min;85 ℃,30 s;4 ℃,永恒。反应体系见表1。
-
反应条件:94 ℃预变性30 s后94 ℃ 10 s,60 ℃ 30 s,共进行40个循环后,进行溶解曲线的检测。反应体系见表2。
表 1 逆转录反应体系
物质 加入量(V/μl) 总RNA X(X=1000/c,c为上述测定的总RNA的浓度) Rnase Free dH2O 16-X 5×Mix 4 表 2 RT-qPCR的反应体系
物质 加入量(V/μl) cDNA 1.0 Primer R 0.3 Primer F 0.3 ddH2O 3.4 SYBR 5.0 总计 10 -
进行Transwell实验以评估细胞在体外的迁移能力。细胞用磷酸缓冲盐水(PBS)洗涤2次,用胰蛋白酶消化,稀释为每毫升含1 × 106个细胞,在Transwell室中,加入200 μl含1% FBS培养基的细胞液,下层用600 μl含10% FBS培养基处理。置于37 ℃、5% CO2的恒温培养箱中培养48 h,然后用棉签小心地擦去未穿过小孔的细胞,再用4%多聚甲醛固定30 min,最后用结晶紫染色1 h。使用倒置显微镜拍摄细胞照片,Image-Pro Plus 6.0分析实验结果。
-
将细胞以5 × 105/孔的比例接种于6孔板中,37 ℃孵育48 h。使用Annexin V-APC/7-AAD凋亡检测试剂盒,按照说明书操作方法,使用流式细胞术检测细胞凋亡。
-
数据分析通过SPSS (21.0)统计软件进行,采用单因素方差分析,实验结果用(
$\bar x$ ± s)表示。与对照组比较,P < 0.05,则认为差异具有统计学意义。 -
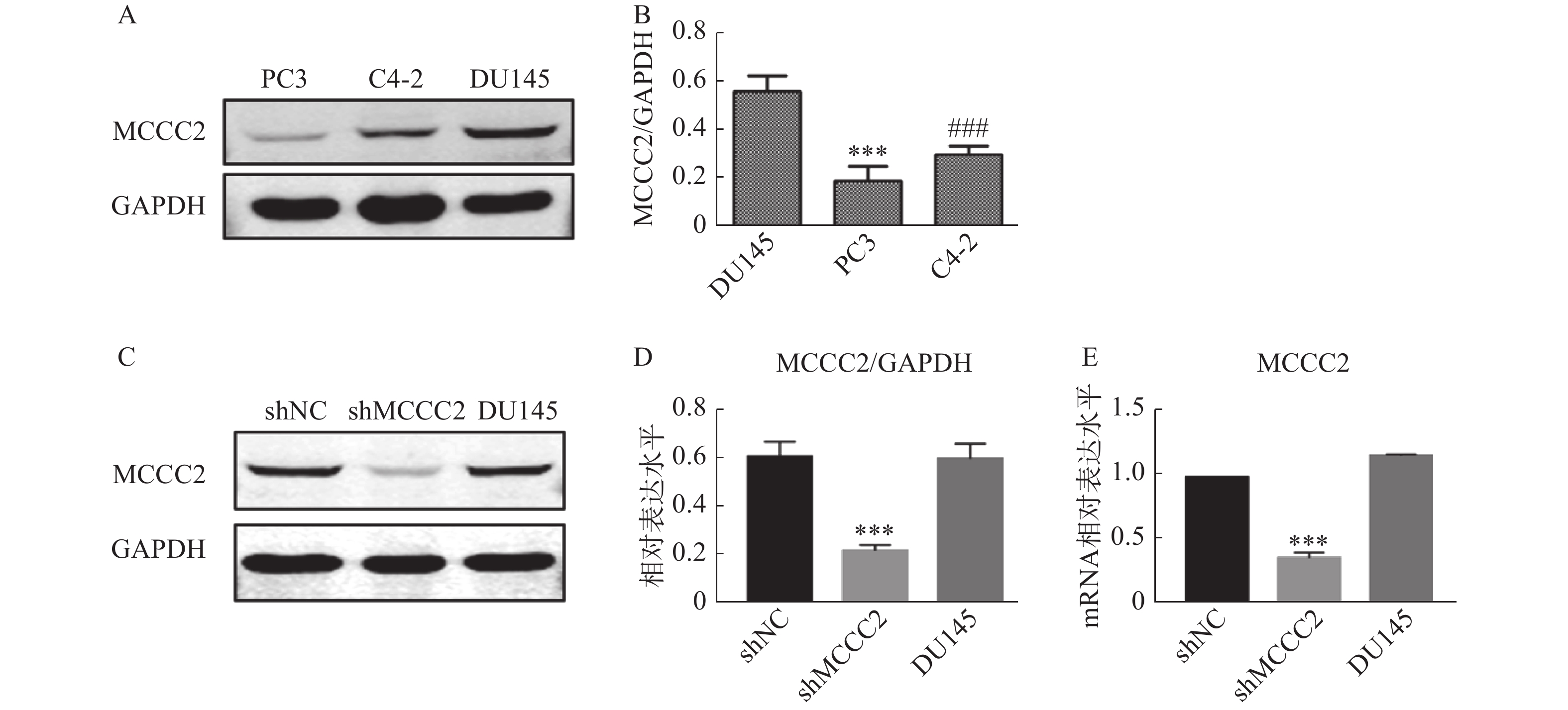
通过Western blot检测PC3、C4-2和DU145细胞中MCCC2的表达水平。如图1A和图1B所示,MCCC2在DU145细胞中的表达为(0.56 ± 0.05);MCCC2在PC3细胞中的表达为(0.24 ± 0.04);MCCC2在C4-2细胞中的表达为(0.32 ± 0.04)。结果表明MCCC2在DU145细胞中的表达水平高于C4-2和PC3细胞。为了探讨MCCC2在前列腺癌发生发展中的生物学作用,我们在DU145细胞中通过慢病毒转染稳定下调了MCCC2的表达。成功建立了低表达MCCC2的DU145细胞株,如图1C和1D,Western blot结果表明,shMCCC2组的MCCC2表达水平明显低于shNC组(0.22 ± 0.02 对 0.61 ± 0.06,P < 0.001),而DU145组与shNC组无明显差异(0.60 ± 0.06 对 0.61 ± 0.06,P > 0.05)。qPCR结果表明,shMCCC2组的MCCC2表达水平明显低于shNC组(0.36 ± 0.04 对 1.0 ±0.00,P < 0.001),而DU145组与shNC组无明显差异(1.17 ± 0.01对1.0 ± 0.00,P > 0.05,图1E)。
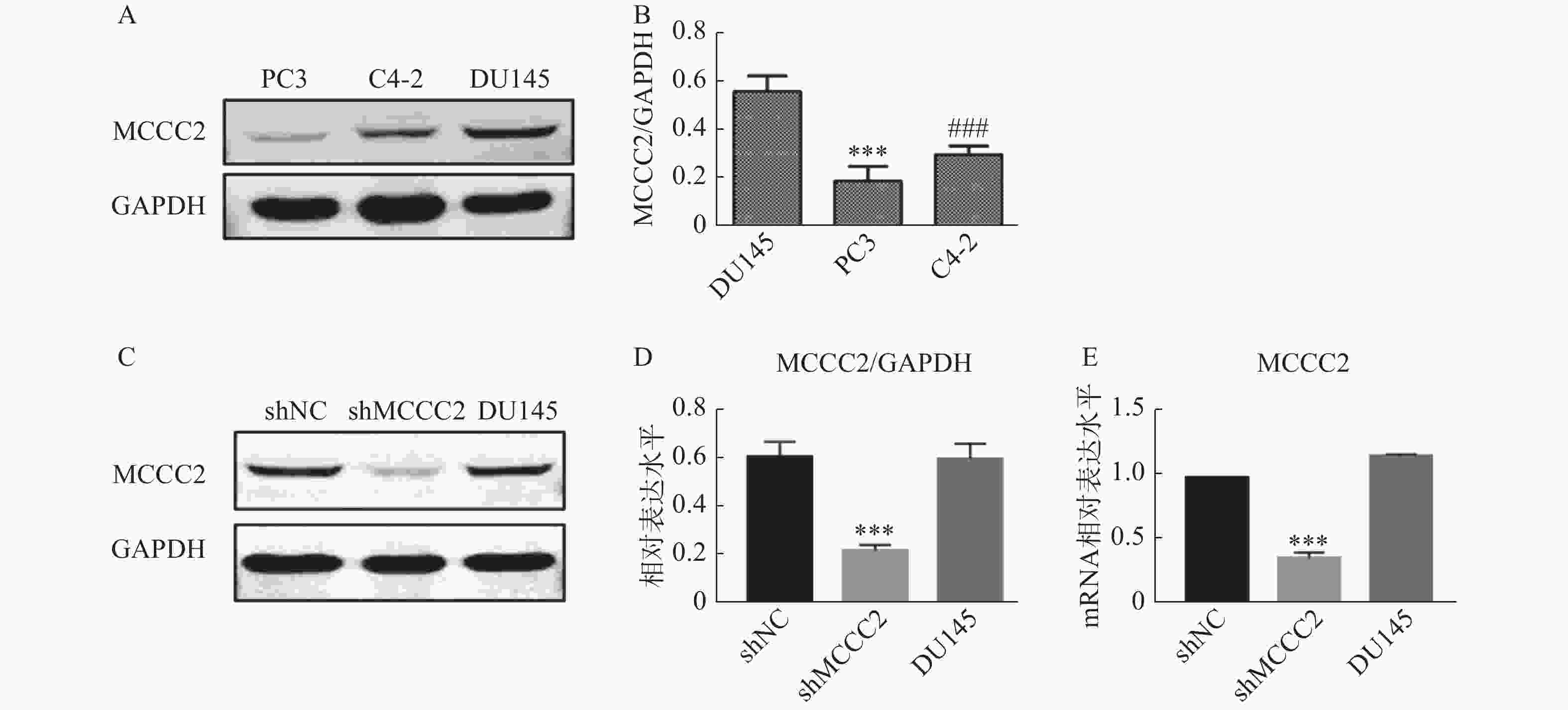
图 1 MCCC2在不同前列腺癌细胞系和shMCCC2 DU145细胞中的表达水平
-
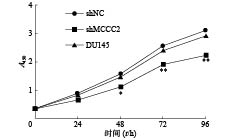
采用CCK-8检测敲减MCCC2对细胞活力的影响。分别在0、24、48、72、96 h后,加CCK-8试剂,在450 nm处测定A值,检测细胞增殖情况。如图2所示,48 h时,shMCCC2组的增殖明显低于shNC组(1.16 ± 0.03 对 1.59 ± 0.07,P < 0.05),而DU145组与shNC组比较无明显差异(1.47 ± 0.05 对 1.59 ± 0.07,P > 0.05)。在72 h时,shMCCC2组的增殖明显低于shNC组(1.92 ± 0.03 对 2.57 ± 0.08,P < 0.01),而DU145组与shNC组比较无明显差异(2.43 ± 0.11对2.57 ± 0.08,P > 0.05)。在96 h时,shMCCC2组的增殖明显低于shNC组(2.24 ±0.04对3.13 ± 0.15,P < 0.01),而DU145组与shNC组比较无明显差异(2.93 ± 0.05对3.13 ± 0.15,P > 0.05)。由结果可以看出,敲减MCCC2能显著抑制细胞增殖,并呈时间依赖性。

图 2 敲减MCCC2对DU145人前列腺癌细胞增殖的影响
-
我们采用Transwell小室实验法测定敲减MCCC2对细胞迁移的影响。采用Image-Pro Plus 6.0分析实验结果,计算细胞迁移率。如图3A和3B所示,shMCCC2组的跨膜DU145细胞数量明显少于shNC组(23.96 ± 1.85对49.73 ± 0.63,P < 0.001);而空白对照组与shNC组相比,跨膜DU145细胞数无显著性差异(48.45 ± 1.26对49.73 ± 0.63,P > 0.05))。由此可见,敲减MCCC2可有效抑制前列腺癌细胞的迁移。

图 3 敲减MCCC2对DU145人前列腺癌细胞迁移能力的影响
-
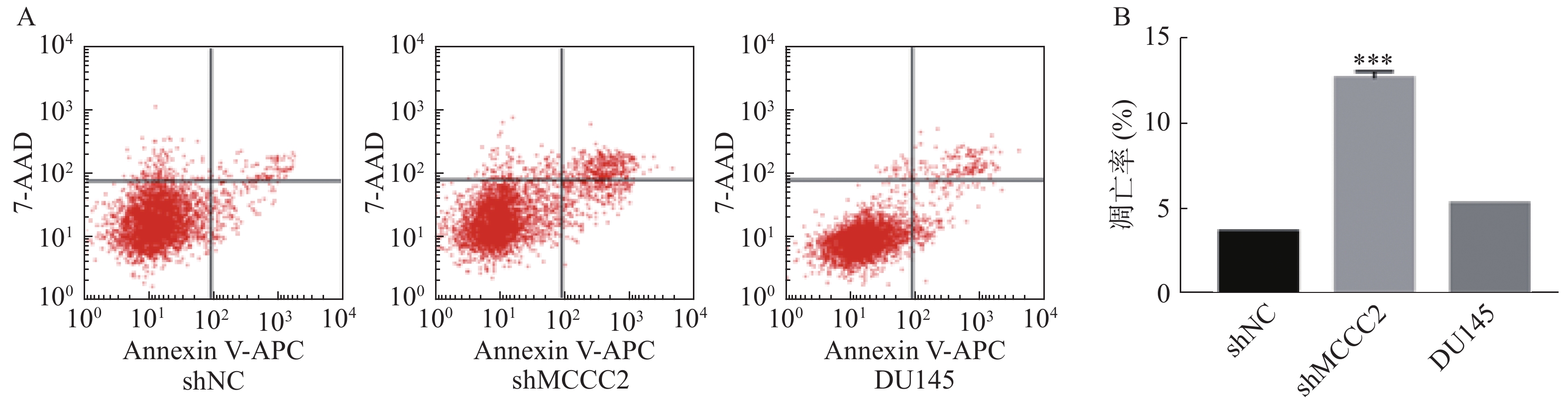
进一步利用Annexin V-APC/7-AAD凋亡检测试剂盒检测了敲减MCCC2对DU145细胞凋亡的影响。结果如图4A和4B所示,shMCCC2组DU145细胞的凋亡率明显高于shNC组(12.64 ± 0.30对 3.68±0.02,P<0.001);空白对照组与shNC组相比,DU145细胞的凋亡率无显著性差异(5.30 ± 0.02对3.68±0.02,P>0.05)。因此,敲减MCCC2可诱导DU145前列腺癌细胞凋亡。
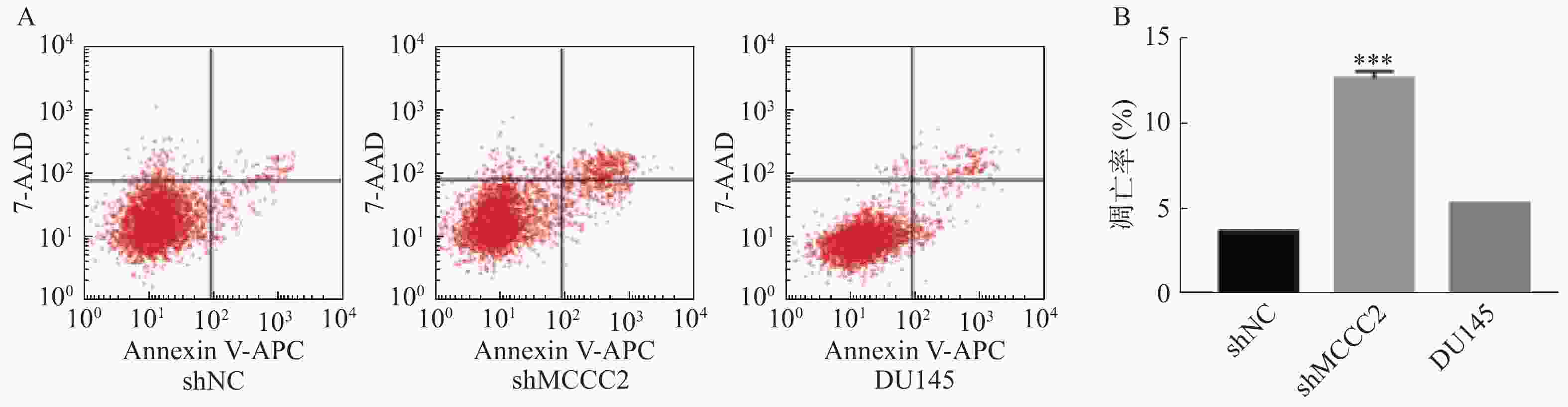
图 4 敲减MCCC2对DU145人前列腺癌细胞凋亡的影响
-
围绕CRPC发生发展中的关键基因和蛋白,已有研究者采用蛋白组学和转录组学等多种组学技术深入挖掘PCa中AR等重要信号通路的关键调节蛋白、基因和潜在的疾病标志物,发现MCCB是前列腺肿瘤组织和正常组织中差异表达的关键蛋白之一[4],而编码该蛋白的基因MCCC2为AR信号通路调节的关键基因,在人类前列腺癌临床样本中显著过表达,可作为潜在的肿瘤标志物[10]。
3-甲基巴豆酰基辅酶A羧化酶(MCC)是一种生物素依赖的羧化酶,可将3-甲基巴豆酰基辅酶A催化为3-甲基戊二酰辅酶A[5-6],是亮氨酸降解途径的重要一步。MCC包含α(MCCA)和β(MCCB)两个亚基,分别由MCCC1和MCCC2基因编码[12-13]。MCC在人体内主要表达于肾脏和肝脏中,它的缺陷可引起甲基巴豆酰甘氨酸尿症,严重的可导致婴儿期的死亡[14-16]。MCC缺陷主要与MCCC1和MCCC2基因的突变有关[17-21]。人MCCC1位于染色体区域3q25-q27,具有19个外显子,而MCCC2位于染色体区域5q12-q13,由17个外显子组成[7-8]。迄今为止,人类基因突变数据库中共报道了MCCC1基因的49个突变和MCCC2基因的52个突变[22]。近年来,越来越多的研究表明,MCCC2参与了包括肝细胞癌、结直肠癌以及乳腺癌在内等多种肿瘤的形成和发展[9-11],其在不同肿瘤中发挥的重要作用有待进一步挖掘。
本研究主要探究了MCCC2与前列腺癌的关系。结果表明,敲减MCCC2可显著抑制DU145前列腺癌细胞的增殖和迁移,并诱导细胞凋亡。研究结果提示,MCCC2可能在前列腺癌发生发展过程中发挥重要作用,有望成为治疗前列腺癌的一个潜在新靶标。至今尚无针对其编码蛋白MCCB的选择性抑制剂的报道,是前列腺癌治疗药物发现的潜在新方向。
Effects of MCCC2 knockdown on proliferation, migration and apoptosis of DU145 prostate cancer cells
-
摘要:
目的 应用慢病毒载体shRNA对DU145细胞中的3-甲基巴豆酰辅酶A羧化酶β亚基编码基因(3-methylcrotonyl-coenzyme A carboxylase 2,MCCC2)进行敲减,探究MCCC2对前列腺癌细胞生物学功能改变的影响。 方法 研究分为3组进行,其中shNC为未敲减MCCC2的DU145细胞对照组,shMCCC2为敲减MCCC2的DU145细胞实验组,DU145为未作任何处理的空白组。运用Western blot和qPCR检测DU145细胞系慢病毒shRNA敲减MCCC2后的表达,CCK8法检测敲减MCCC2对DU145细胞增殖的影响;Transwell检测敲减MCCC2对DU145细胞迁移的影响;流式细胞术检测敲减MCCC2对DU145细胞凋亡的影响。 结果 前列腺癌细胞系DU145经慢病毒shRNA敲减MCCC2后,shMCCC2组的MCCC2表达水平明显低于shNC组(0.22 ± 0.02对0.61 ± 0.06,P < 0.001),shMCCC2组增殖(2.24 ± 0.04对3.13 ± 0.15)和迁移(23.96 ± 1.85对49.73 ± 0.63)均明显低于shNC组(P<0.001)、而shMCCC2组凋亡(12.64 ± 0.30对3.68 ± 0.02)明显高于shNC组(P<0.001)。 结论 敲减MCCC2可显著抑制DU145细胞的增殖和迁移并诱导细胞凋亡,提示MCCC2下调与DU145细胞系肿瘤生物学特性的改变具有一定相关性,有望成为前列腺癌的潜在治疗靶标。 Abstract:Objective To investigate the change of biological characteristics after stable knockdown of the coding gene of 3-methylcrotonyl-coenzyme A carboxylase β subunit (MCCC2) expression in DU145 by lentivirus shRNA. Methods Three groups were included in this study. shNC was the control group in which MCCC2 was negatively knocked down in DU145. shMCCC2 was the experimental group in which MCCC2 was knocked down. DU145 was the blank group without any treatment. The expression of MCCC2 was assessed by Western blot and qPCR. The proliferation of DU145 cells was detected by CCK8 assay. The migration ability of DU145 was detected by transwell. The apoptosis of DU145 cells was detected by flow cytometry. Results The expression level of MCCC2 in shMCCC2 group was significantly lower than that in shNC group (0.22 ± 0.02 vs 0.61 ± 0.06, P < 0.001). The proliferation (2.24 ± 0.04 vs 3.13 ± 0.15) and migration (23.96 ± 1.85 vs 49.73 ± 0.63) of DU145 cells in shMCCC2 group was significantly lower than that in shNC group, whereas the apoptosis (12.64 ± 0.30 vs 3.68 ± 0.02) of DU145 cells in shMCCC2 was significantly higher than that in shNC group. Conclusion MCCC2 knockdown significantly inhibited the proliferation and migration, and induced apoptosis of DU145 cells, which indicated that the down-regulation of MCCC2 is correlated with the change of tumor biological characteristics of DU145 cell line and can be a potential target for the treatment of prostate cancer. -
Key words:
- MCCC2 /
- prostate cancer /
- proliferation /
- migration /
- apoptosis
-
图 1 MCCC2在不同前列腺癌细胞系和shMCCC2 DU145细胞中的表达水平
A.Western blot法检测PC3、C4-2和DU145细胞中MCCC2的蛋白表达水平;B.MCCC2蛋白表达水平的灰度值分析,***P < 0.001,与DU145比较;###P < 0.001,与DU145比较;C.Western blot法检测shMCCC2 DU145细胞中MCCC2的蛋白表达水平;D.不同处理组细胞MCCC2蛋白表达水平的灰度值分析,***P < 0.001,与shNC比较;E.RT-qPCR检测shMCCC2 DU145细胞中MCCC2的mRNA表达水平,***P < 0.001,与shNC比较。
图 3 敲减MCCC2对DU145人前列腺癌细胞迁移能力的影响
A.200倍镜下不同处理组的细胞迁移图;B.Image-Pro Plus 6.0分析不同处理组的细胞迁移面积,***P < 0.001,与shNC比较。
-
[1] HA CHUNG B, HORIE S, CHIONG E. The incidence, mortality, and risk factors of prostate cancer in Asian men[J]. Prostate Int,2019,7(1):1-8. doi: 10.1016/j.prnil.2018.11.001 [2] 唐志柳, 白洁, 顾丽娜, 等. 2000-2010年我国前列腺癌和乳腺癌流行状况的系统性综述[J]. 中国肿瘤, 2013, 22(04):260-265. [3] 王宁, 刘硕, 杨雷, 等. 2018全球癌症统计报告解读[J]. 肿瘤综合治疗电子杂志, 2019, 5(01):87-97. [4] ABULA A, SHAO W, TUSONG H, et al. Protein expression information of prostate infection based on data mining[J]. J Infect Public Health,2020,13(10):1533-1536. doi: 10.1016/j.jiph.2019.07.019 [5] GRÜNERT S C, STUCKI M, MORSCHER R J, et al. 3-methylcrotonyl-CoA carboxylase deficiency: clinical, biochemical, enzymatic and molecular studies in 88 individuals[J]. Orphanet J Rare Dis,2012,7:31. doi: 10.1186/1750-1172-7-31 [6] MORSCHER R J, GRÜNERT S C, BÜRER C, et al. A single mutation in MCCC1 or MCCC2 as a potential cause of positive screening for 3-methylcrotonyl-CoA carboxylase deficiency[J]. Mol Genet Metab,2012,105(4):602-606. doi: 10.1016/j.ymgme.2011.12.018 [7] HOLZINGER A, RÖSCHINGER W, LAGLER F, et al. Cloning of the human MCCA and MCCB genes and mutations therein reveal the molecular cause of 3-methylcrotonyl-CoA: carboxylase deficiency[J]. Hum Mol Genet,2001,10(12):1299-1306. doi: 10.1093/hmg/10.12.1299 [8] GALLARDO M E, DESVIAT L R, RODRÍGUEZ J M, et al. The molecular basis of 3-methylcrotonylglycinuria, a disorder of leucine catabolism[J]. Am J Hum Genet,2001,68(2):334-346. doi: 10.1086/318202 [9] PANG J, LIU W P, LIU X P, et al. Profiling protein markers associated with lymph node metastasis in prostate cancer by DIGE-based proteomics analysis[J]. J Proteome Res,2010,9(1):216-226. doi: 10.1021/pr900953s [10] MARQUES R B, DITS N F, ERKENS-SCHULZE S, et al. Modulation of androgen receptor signaling in hormonal therapy-resistant prostate cancer cell lines[J]. PLoS One,2011,6(8):e23144. doi: 10.1371/journal.pone.0023144 [11] LIU Y, YUAN Z, SONG C. Methylcrotonoyl-CoA carboxylase 2 overexpression predicts an unfavorable prognosis and promotes cell proliferation in breast cancer[J]. Biomarks Med,2019,13(6):427-436. doi: 10.2217/bmm-2018-0475 [12] JUNG C W, LEE B H, KIM J H, et al. Uneventful clinical courses of Korean patients with methylcrotonylglycinuria and their common mutations[J]. J Hum Genet,2012,57(1):62-64. doi: 10.1038/jhg.2011.116 [13] AL-SHAMSI A, HERTECANT J L, SOUID A K, et al. Whole exome sequencing diagnosis of inborn errors of metabolism and other disorders in United Arab Emirates[J]. Orphanet J Rare Dis,2016,11(1):94. doi: 10.1186/s13023-016-0474-3 [14] RIPS J, ALMASHANU S, MANDEL H, et al. Primary and maternal 3-methylcrotonyl-CoA carboxylase deficiency: insights from the Israel newborn screening program[J]. J Inherit Metab Dis,2016,39(2):211-217. doi: 10.1007/s10545-015-9899-4 [15] ZANDBERG L, VAN DYK H C, VAN DER WESTHUIZEN F H, et al. A 3-methylcrotonyl-CoA carboxylase deficient human skin fibroblast transcriptome reveals underlying mitochondrial dysfunction and oxidative stress[J]. Int J Biochem Cell Biol,2016,78:116-129. doi: 10.1016/j.biocel.2016.07.010 [16] FICICIOGLU C, PAYAN I. 3-methylcrotonyl-CoA carboxylase deficiency: metabolic decompensation in a noncompliant child detected through newborn screening[J]. Pediatrics,2006,118(6):2555-2556. doi: 10.1542/peds.2006-1659 [17] BAUMGARTNER M R, ALMASHANU S, SUORMALA T, et al. The molecular basis of human 3-methylcrotonyl-CoA carboxylase deficiency[J]. J Clin Invest,2001,107(4):495-504. doi: 10.1172/JCI11948 [18] STADLER S C, POLANETZ R, MAIER E M, et al. Newborn screening for 3-methylcrotonyl-CoA carboxylase deficiency: population heterogeneity of MCCA and MCCB mutations and impact on risk assessment[J]. Hum Mutat,2006,27(8):748-759. doi: 10.1002/humu.20349 [19] YANG L, YANG J, ZHANG T, et al. Identification of eight novel mutations and transcript analysis of two splicing mutations in Chinese newborns with MCC deficiency[J]. Clin Genet,2015,88(5):484-488. doi: 10.1111/cge.12535 [20] FONSECA H, AZEVEDO L, SERRANO C, et al. 3-methylcrotonyl-CoA carboxylase deficiency: mutational spectrum derived from comprehensive newborn screening[J]. Gene,2016,594(2):203-210. doi: 10.1016/j.gene.2016.09.003 [21] COZZOLINO C, VILLANI G R, FRISSO G, et al. Biochemical and molecular characterization of 3-methylcrotonylglycinuria in an Italian asymptomatic girl[J]. Genet Mol Biol,2018,41(2):379-385. doi: 10.1590/1678-4685-gmb-2017-0093 [22] STENSON P D, MORT M, BALL E V, et al. The Human Gene Mutation Database: building a comprehensive mutation repository for clinical and molecular genetics, diagnostic testing and personalized genomic medicine[J]. Hum Genet,2014,133(1):1-9. doi: 10.1007/s00439-013-1358-4 -
 点击查看大图
点击查看大图
图(4) / 表(2)
计量
- 文章访问数: 2216
- HTML全文浏览量: 717
- PDF下载量: 18
- 被引次数: 0

